7 Microbial Regulatory Systems
I DNA-Binding Proteins and Transcriptional Regulation
As Bacterial Cells Chatter, Viruses Eavesdrop
Despite their primitive nature, bacterial and archaeal cells participate in a form of communication called quorum sensing. Quorum sensing coordinates behaviors that require a certain cell density to be successful, such as virulence and biofilm formation. This communication is dependent on the microbial secretion of autoinducer molecules. As the population of autoinducer-producing cells increases in a given environment, so does the local concentration of autoinducer. Once the autoinducer reaches a certain threshold, regulatory proteins respond by activating genes whose products are necessary for group behavior.
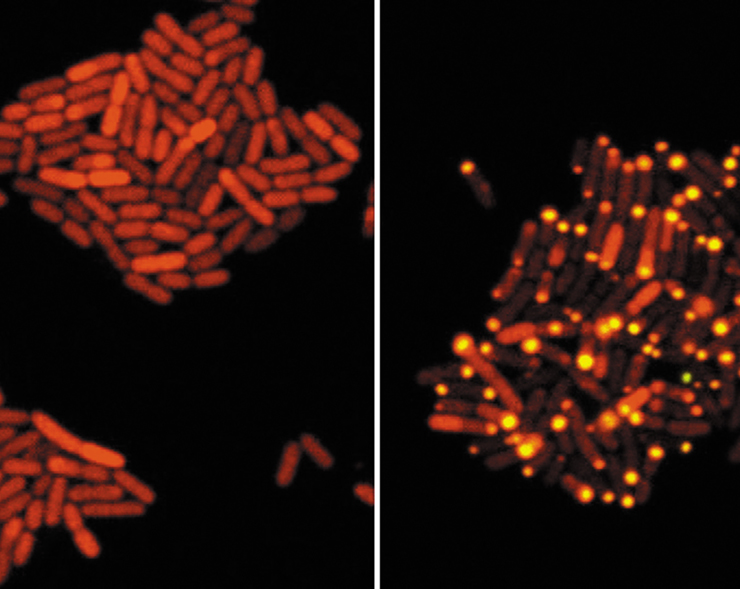
As extensive research on quorum sensing continued, microbiologists began to ponder if viruses could respond to quorum sensing. Temperate bacteriophages have two possible “lifestyles”—their DNA can reside in dormancy within the bacterial host cell chromosome or they can aggressively multiply and lyse their host cell. Using elegant experiments, researchers discovered that certain dormant bacteriophages are actually “listening in” on their hosts’ conversation. This eavesdropping occurs through a phage-encoded receptor protein that can sense host autoinducer concentrations. In response to autoinducer concentrations, this phage regulatory protein initiates a cascade of events that awakens the phage from dormancy.
The image to the left shows Escherichia coli cells harboring a fluorescently labeled phage repressor protein called cI that is distributed uniformly throughout the host cell and is essential for phage dormancy. The panel on the right shows the same cells with the addition of the newly discovered phage-encoded protein called Qtip, which is only produced in response to high autoinducer levels. Qtip functions by inactivating the cI protein by aggregating it (bright foci in right panel). Without free cI, the dormant phage produces thousands of progeny and lyses its host. This fascinating phenomenon illustrates how a nonliving microbe—the virus—both senses and responds to host cell communication to ensure its own successful spread!
Source: Silpe, J.E., and B.L. Bassler. 2019. A host-produced quorum-sensing autoinducer controls a phage lysis-lysogeny decision. Cell 176: 268.
To efficiently orchestrate the numerous reactions that occur in a cell and to maximally use available resources, cells must regulate the types, amounts, and activities of proteins and other macromolecules they make. Some proteins and RNA molecules are needed in the cell at about the same level under all growth conditions, and the production of these molecules is said to be constitutive. However, many proteins and RNAs are needed under some conditions but not others. For instance, enzymes required to catabolize the sugar lactose are useful only if lactose is available to the cell. Regulatory systems can thus be viewed as a means to improve the overall fitness of an organism and maximize its ability to produce offspring from the resources available.
Microbes regulate protein function in two distinct ways. One mechanism controls the amount of an enzyme or other protein, whereas the second controls the activity of a preformed enzyme or other protein (Figure 7.1). The amount of protein synthesized can be regulated at either the level of transcription—by varying the amount of mRNA made—or the level of translation—by translating or not translating the mRNA. Collectively, these regulatory processes are called gene expression. After a protein has been synthesized, post-translational regulatory processes can further regulate the activity of some proteins (Figure 7.1). In this chapter we focus on how the cell efficiently controls its metabolism by regulating both enzyme synthesis and enzyme activity.
Figure 7.1 Gene expression and regulation of protein activity.

For DNA, the promoter and terminator as well as regions related to transcriptional activation and repression are indicated. Cellular signals control the activity and DNA-binding ability of activator and repressor proteins. For mRNA, the 5′ untranslated region (5′-UTR) is a short region between the start of transcription and the start of translation, including the ribosome-binding site (RBS). The 3′ untranslated region (3′-UTR) is a short region between the stop codon and the transcription terminator. These are the regions where translational regulation often occurs, with riboswitches and small RNAs (sRNAs) often targeting the 5′-UTR in response to cellular signals. Mechanisms for regulating protein activity after translation are shown near the bottom.
I: DNA-Binding Proteins and Transcriptional Regulation
I: DNA-Binding Proteins and Transcriptional Regulation
I DNA-Binding Proteins and Transcriptional Regulation
**Transcriptional controls in Bacteria and Archaea typically function by binding a specific regulatory protein to a specific region of DNA, an event that either represses or activates transcription.**
Although several microbial regulatory mechanisms are possible, our discussion begins with control at the level of transcription because this is the major means of regulation in prokaryotic cells.
7.1 DNA-Binding Proteins
Before delving into the themes of transcriptional regulation, we must first recall our discussion of how bacterial and archaeal gene arrangement differs from that of eukaryotes (Chapter 6). Besides lacking traditional introns, bacterial and archaeal genes can be arranged in operons—two or more genes transcribed under the control of a single regulatory site (Section 6.5). This regulatory site, or promoter region, is located upstream of where RNA polymerase initiates transcription (Figure 7.1). Bacterial and archaeal promoter regions are characterized by distinct nucleotide sequences that must be recognized and bound by DNA-binding proteins for RNA polymerase to bind and allow transcription to occur (Sections 6.5 and 6.6).
Interaction of Proteins with Nucleic Acids
Interactions between proteins and nucleic acids are central to replication, transcription, and translation, and also to the regulation of these processes. Protein–nucleic acid interactions may be specific or nonspecific, depending on whether the protein attaches anywhere along the nucleic acid or binds to a specific site. Most DNA-binding proteins interact with DNA in a sequence-specific manner. Specificity is provided by interactions between specific amino acid side chains of the proteins and specific chemical groups on the nitrogenous bases and the sugar–phosphate backbone of the DNA. Because of its size, the major groove of DNA (Figure 6.3) is the main site of protein binding, and Figure 6.1c identified atoms of the bases in the major groove that are known to interact with proteins. To achieve high specificity, the binding protein must interact simultaneously with several nucleotides.
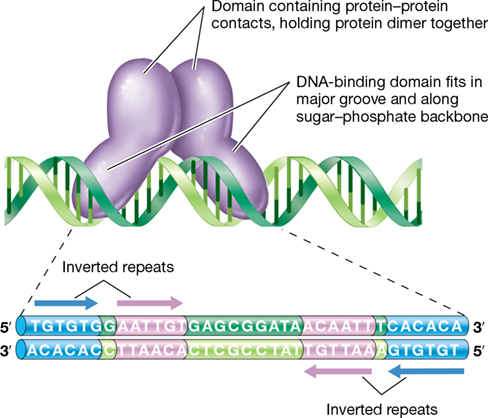
In DNA, an inverted repeat is a nucleotide sequence followed downstream by its inverted complement (Section 6.5). Such inverted repeats are frequently the locations at which proteins bind specifically to DNA (Figure 7.2). Note that this interaction does not require the formation of stem–loop structures in the DNA as was described for the signal for termination of transcription (Figure 6.23). DNA-binding proteins are often homodimeric, meaning they are composed of two identical polypeptide subunits, each subdivided into domains—regions of the protein with a specific structure and function. Each subunit has a domain that interacts specifically with a region of DNA in the major groove. When protein dimers interact with inverted repeats on DNA, each subunit binds to one of the inverted repeats. The dimer as a whole thus binds to both DNA strands (Figure 7.2).
Figure 7.2 DNA-binding proteins.

Many DNA-binding proteins are dimers that combine specifically with two sites on the DNA. The specific DNA sequences that interact with the protein are inverted repeats. The nucleotide sequence of the operator gene of the lactose operon (see Figure 7.6) is shown, and the inverted repeats, which are sites at which the lac repressor (LacI) makes contact with the DNA, are shown in purple and blue boxes.
Structure of DNA-Binding Proteins
DNA-binding proteins in both Bacteria and Archaea as well as eukaryotes possess several classes of protein domains that are critical for proper binding to DNA. One of the most common is the helix-turn-helix structure (**Figure 7.3*a***). This consists of two segments of polypeptide chain that have an α-helix secondary structure connected by a short sequence forming the “turn.” The first helix is the recognition helix, which interacts specifically with DNA. The second helix, the stabilizing helix, stabilizes the first helix by interacting with it by way of hydrophobic interactions. The turn linking the two helices consists of three amino acid residues, the first of which is typically a glycine. Sequences are recognized by noncovalent interactions, including hydrogen bonds and van der Waals contacts, between the recognition helix of the protein and specific chemical groups in the sequence of base pairs on the DNA.
Figure 7.3 The helix-turn-helix structure of some DNA-binding proteins.

(a) A simple model of the helix-turn-helix structure within a single protein subunit. (b) A computer model of both subunits of the bacteriophage lambda repressor bound to its operator. The complementary DNA strands are red and blue. One subunit of the dimeric repressor protein is shown in orange and the other in yellow. Each subunit contains a helix-turn-helix structure. Inset: Atomic force microscopy showing copies of the LacI repressor protein (arrow) bound to multiple operator sites on a DNA molecule.
Many different DNA-binding proteins from Bacteria contain the helix-turn-helix structure. These include many repressor proteins, such as the lac and trp repressors of Escherichia coli (Section 7.3 and Figure 7.3 inset), and some proteins of bacterial viruses, such as the bacteriophage lambda repressor (Figure 7.3b). Indeed, over 250 different proteins with this motif bind to specific locations on DNA to regulate transcription in E. coli. Two other types of protein domains are commonly found in DNA-binding proteins. One of these, the zinc finger, is frequently found in regulatory proteins in eukaryotes and, as its name implies, binds a zinc ion. The other protein domain commonly found in DNA-binding proteins is the leucine zipper, which contains regularly spaced leucine residues that function to hold two recognition helices in the correct orientation to bind DNA.
Once a protein binds at a specific site on the DNA, several outcomes are possible. Some DNA-binding proteins are enzymes that catalyze a specific reaction on the DNA, such as transcription. In other cases, however, the binding event either blocks transcription or activates it, and we consider these now.
Check Your Understanding
Why are most DNA-binding proteins specific to certain chemical groups within the DNA?
7.2 Transcription Factors and Effectors
Transcription is the first step in biological information flow; because of this, it is simple and efficient to control gene expression at this point. If one gene is transcribed more frequently than another, there will be more of its mRNA available for translation and therefore a greater amount of its protein product in the cell. This control is mediated by transcription factors and effector molecules.
Mechanisms of Transcription Factors
Proteins that control the rate of gene transcription by binding to specific DNA sequences are called transcription factors. Depending on the type of transcription factor, this DNA binding can lead to increased or decreased transcription of the associated gene. If binding to the DNA results in turning transcription on, the transcription factor is called an activator protein (**Figure 7.4*a***). Conversely, binding of a repressor protein to DNA results in turning gene expression off (Figure 7.4b).
Figure 7.4 Transcription factors, effectors, and allosterism.

Binding of effector molecules to activator and repressor proteins results in an allosteric change that affects the DNA-binding ability of the transcription factors. (a) Binding of an activator to the DNA results in recruiting RNA polymerase and turning transcription on. (b) Binding of a repressor protein to the operator region of the DNA results in blocking RNA polymerase and turning transcription off.
Activator proteins turn on transcription by binding to DNA and helping to recruit RNA polymerase, or a sigma (σ) factor in the case of bacterial transcription (Section 6.5), to a specific promoter region (Figure 7.4a). Repressor proteins turn off transcription by binding to a specific DNA sequence of a gene called an operator region (Figure 7.4b). This region gets its name from the operon, a cluster of consecutive genes whose expression is under the control of a single operator (Section 6.2). All of the genes in an operon are transcribed as a single unit yielding a single mRNA (Section 6.5). The operator is located downstream of the promoter where synthesis of mRNA is initiated (Figure 7.4). The activity of activator and repressor proteins is described in more detail in the next section.
Effector Molecules
The ability of activator and repressor proteins to bind DNA is controlled by small molecules called effectors. Transcription factors are allosteric proteins; that is, their conformation is altered when the effector molecule binds to them (Figure 7.4 and Section 7.15). This conformational change determines if the transcription factor can bind DNA. If the binding of an effector to a transcription factor results in turning transcription on, the molecule is considered an inducer (Figure 7.4a). Conversely, corepressor molecules are those whose binding to a transcription factor results in turning transcription off (Figure 7.4b).
Effector molecules are typically cell metabolites, such as substrates or end products of the pathways that they regulate. As we will discuss with the lac operon—the operon encoding genes of lactose catabolism—structural analogs of effector molecules can also control transcription factor binding even though they are not themselves substrates for the enzymes that the operon encodes (Section 7.8). Thus, small molecules can indirectly regulate gene expression by binding to transcription factors, as many examples in this chapter will demonstrate.
Check Your Understanding
What is a transcription factor and what are the two major types?
What role do effectors have on transcription factors?
7.3 Repression and Activation
The activities of activator and repressor proteins are simple forms of regulation that govern gene expression at the level of transcription, and the binding of these transcription factors to DNA plays the initial role in controlling the kinds and activities of enzymes in a cell. We begin our discussion of the activities of some common transcription factors by considering cellular conditions in which the expression of particular sets of enzymes are controlled.
Mastering Microbiology
Activities: Microbiology Animation: Operons: Repression
Enzyme Repression and Induction
Often the enzymes that catalyze the synthesis of a specific product are not made if the product is already present in the growth medium in sufficient amounts. For example, in Escherichia coli and many other Bacteria, the enzymes needed to synthesize the amino acid arginine (Figure 6.27) are made only when arginine is absent from the culture medium; an excess of arginine decreases the synthesis of these enzymes. This is called enzyme repression.
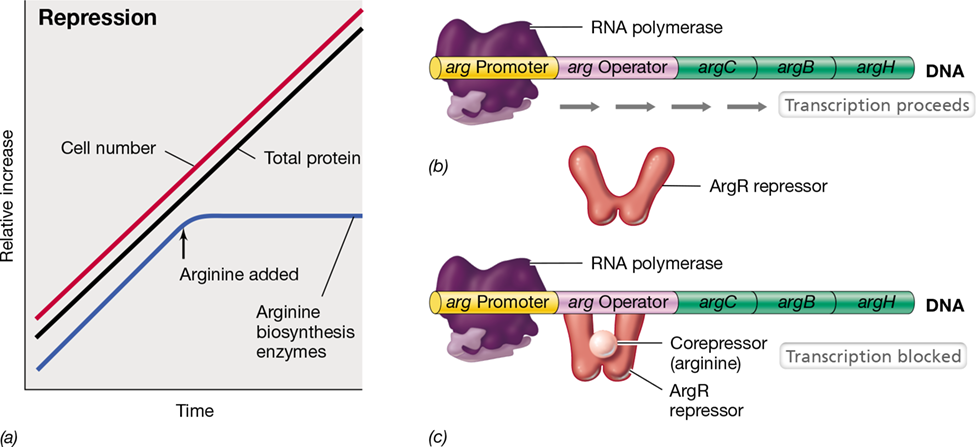
As can be seen in **Figure 7.5*a***, if arginine is added to a culture growing exponentially in a medium devoid of arginine, growth continues at the previous rate, but production of the enzymes for arginine synthesis stops. Note that this is a specific effect, as the synthesis of all other enzymes in the cell continues at the previous rate. This is because the enzymes affected by a particular repression event make up only a tiny fraction of the entire complement of proteins in the cell. Enzyme repression is widespread in Bacteria as a means of controlling the synthesis of enzymes required for the production of amino acids and the nucleotide precursor purines and pyrimidines (Section 3.14). In most cases, the final product of a particular biosynthetic pathway is the effector molecule (corepressor) for that pathway and must be present in excess of biosynthetic needs to repress the enzymes of the pathway. Such enzyme repression ensures that the organism does not waste energy and nutrients synthesizing unneeded enzymes.
Figure 7.5 Enzyme repression and expression of the arginine operon.

(a) In a growing bacterial culture, the addition of arginine to the medium specifically represses production of enzymes needed to make arginine. Net protein synthesis is unaffected. (b) The operon is transcribed because the repressor ArgR is unable to bind to the operator. (c) After the corepressor for the argCBH operon, the amino acid arginine, binds to the repressor, the repressor binds to the operator and blocks transcription; mRNA and the proteins it encodes are not made.
Enzyme induction is conceptually the opposite of enzyme repression. In enzyme induction, an enzyme is made only when its substrate is present. Enzyme repression typically affects biosynthetic (anabolic) enzymes. By contrast, enzyme induction usually affects the synthesis of specific degradative (catabolic) enzymes. To illustrate induction, consider the utilization of the sugar lactose as a carbon and energy source by E. coli. The enzymes for using lactose are encoded by the lac operon (Figure 7.6). Figure 7.6a shows the induction of β-galactosidase, the enzyme that cleaves lactose into glucose and galactose. This enzyme is required for E. coli to grow on lactose. If lactose is absent, the enzyme is not made, but synthesis begins almost immediately after lactose is added. The three genes in the lac operon encode three proteins, including β-galactosidase, that are induced simultaneously upon adding lactose. Such enzyme induction ensures that specific enzymes are synthesized only when needed.
Figure 7.6 Enzyme induction and expression of the lactose operon.

(a) In a growing bacterial culture, the addition of lactose to the medium specifically induces synthesis of the enzyme β-galactosidase and other proteins of lactose catabolism. Net protein synthesis is unaffected. (b) Binding of the LacI repressor protein to the operator blocks the activity of RNA polymerase. (c) Binding of the inducer molecule to the LacI repressor inactivates LacI so that it no longer can bind to the operator. This type of inducer binding results in derepression because RNA polymerase can now transcribe an mRNA for that operon. For the lac operon, the sugar allolactose rather than lactose is the actual inducer that binds to the LacI repressor.
Mechanisms of Repression and Derepression
As discussed in Section 7.2, repressor proteins that bind to DNA turn off transcription, and if a repressor binds DNA only in the presence of its effector, the effector is considered a corepressor. Figure 7.5b shows transcription of the arginine genes, which proceeds when the cell needs arginine. However, when arginine is plentiful, it becomes a corepressor. As Figure 7.5c shows, arginine binds to a specific repressor protein, the arginine repressor (ArgR), present in the cell. Binding of the corepressor to ArgR results in an allosteric change in this protein that allows it to bind to the operator. In this way, transcription is physically blocked because RNA polymerase can neither bind nor proceed, and in the absence of arg mRNA, the corresponding polypeptides encoded by the arg operon cannot be synthesized. If the mRNA is polycistronic (Section 6.5), as the arg operon is, all the polypeptides encoded by this mRNA will be repressed (Figure 7.5c).
In contrast to the control of the arg operon, some repressor proteins actively bind to operator regions on the DNA in the absence of their effector molecule. Figure 7.6b shows the repression of the lac operon—an example of an inducible operon—by the binding of the lactose repressor (LacI) to the operator region. This binding of LacI completely blocks transcription. If the LacI effector molecule is present, it combines with the repressor protein, causing an allosteric change that prevents LacI from binding DNA; when this occurs, inhibition is overcome and transcription can proceed (Figure 7.6c). Because control of inducible operons is the opposite of repression, it is also called derepression. The corresponding effector molecule (corepressor) in this case is called an inducer as its presence leads to inactivation of the repressor and ultimately enzyme synthesis.
Mastering Microbiology
Activities: Microbiology Animation: Operons: Induction
For the lac operon example in Figure 7.6c, allolactose, an isomer made from lactose, is the actual inducer molecule that leads to the expression of the lac operon and ultimately synthesis of the enzyme β-galactosidase (encoded by lacZ). However, isopropylthiogalactoside (IPTG), a nonmetabolizable analog of allolactose that also induces the lac operon, is often used in the laboratory for detailed studies of lactose utilization and β-galactosidase activity in E. coli.
All regulatory systems employing repressors have the same underlying mechanism: prevention of mRNA synthesis by the activity of specific repressor proteins that are themselves under the control of specific small effector molecules. And, as previously noted, because the repressor’s role is to stop transcription, regulation by repressors is called negative control. Another point to note is that genes are not turned on and off completely like light switches. DNA-binding proteins vary in concentration and affinity and thus control is quantitative. Even when a gene is “fully repressed” there is often a very low level of basal transcription.
Mechanisms of Activation
In contrast to repressor proteins binding and preventing transcription of DNA, some operons are transcribed only if a specific activator protein is first bound to the DNA (Figure 7.7). Activator protein binding is necessary in these cases because the promoter sequences in these operons are poor matches to consensus promoter sequences (Section 6.5) and hence bind RNA polymerase only weakly. Thus, even with the correct sigma (σ) factor, the RNA polymerase has difficulty recognizing these promoters. And, in contrast to the case where operons are under repressor control (Figures 7.5 and 7.6), genes controlled by activator proteins are under positive control because the role of activator proteins is to facilitate rather than block transcription.
Figure 7.7 Activator protein interactions with RNA polymerase.

(a) The activator-binding site is near the promoter. (b) The activator-binding site is several hundred base pairs from the promoter. In this case, the DNA must be looped to allow the activator and the RNA polymerase to interact.
The function of activator proteins is to help the RNA polymerase recognize the promoter and begin transcription. For example, the activator protein may modify the structure of the DNA by bending it (Figure 7.8), allowing the RNA polymerase to make necessary contacts with nucleotides in the promoter region to begin transcription. Alternatively, the activator protein may interact directly with the RNA polymerase. This can happen either when the activator-binding site is close to the promoter (Figure 7.7a) or when it is several hundred base pairs away from the promoter, a situation in which DNA looping is required to make the necessary contacts between protein and nucleic acid (Figure 7.7b).
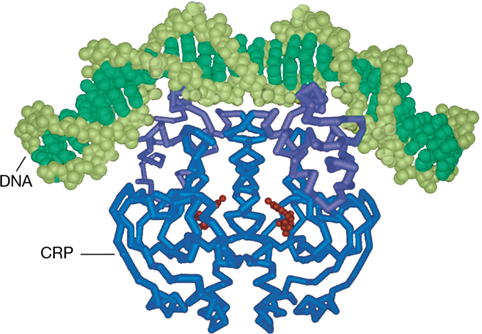
Figure 7.8 Computer model of a positive regulatory protein interacting with DNA.

This model shows the cyclic AMP receptor protein (CRP), a regulatory protein that controls several operons. The α-carbon backbone of this protein is shown in blue and purple, while residues that bind cyclic AMP are shown in red. The protein is binding to a DNA double helix (shades of green). Note that binding of the CRP protein to DNA has bent the DNA.
An example of activation is the regulation of the maltose operon in Escherichia coli, which encodes the enzymes necessary for the catabolism of the disaccharide sugar maltose. This operon possesses a maltose activator protein called MalT that cannot bind to the DNA unless it first binds to its effector molecule, maltose (Figure 7.9). Thus, maltose is also considered an inducer molecule because the enzymes for maltose catabolism in E. coli are synthesized only after the addition of maltose to the medium. This follows the pattern shown for β-galactosidase in Figure 7.6c except that maltose rather than lactose is required to induce gene expression. When the maltose activator protein binds to DNA, it allows RNA polymerase to begin transcription (Figure 7.9b). Like repressor proteins, activator proteins bind specifically to particular sites within the DNA. However, the region on the DNA that is the binding site of the activator is not called an operator (Figures 7.5 and 7.6) but instead an activator-binding site (Figures 7.7 and 7.9). Nevertheless, the genes controlled by this activator-binding site are still called an operon.
Figure 7.9 Positive control of enzyme induction in the maltose operon.

(a) In the absence of an inducer, neither the MalT activator protein nor the RNA polymerase can bind to the DNA. (b) When the sugar maltose (the inducer of the malEFG operon) is present, it binds to the MalT activator protein, which in turn binds to the activator-binding site. This recruits RNA polymerase to bind to the mal promoter and begin transcription.
Operons versus Regulons
In E. coli, the genes required for maltose utilization are spread out over the chromosome in several operons, each of which has an activator-binding site to which a copy of the maltose activator protein can bind (Figure 7.10). Therefore, the maltose activator protein actually controls the transcription of more than one operon. When more than one operon is under the control of a single regulatory protein, these operons are collectively called a regulon. Thus, the enzymes for maltose utilization are encoded by the maltose regulon. A set of operons whose promoters are recognized only by a specific alternative sigma (σ) factor (Table 6.3) is another example of a positively regulated regulon.
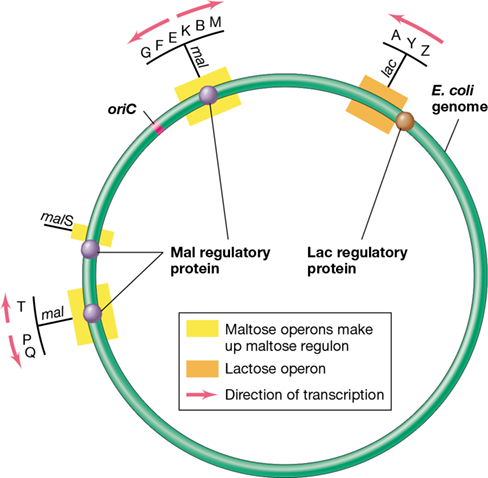
Figure 7.10 Maltose regulon of *Escherichia coli*.

The genes and operons required for maltose utilization (mal) are dispersed throughout the E. coli genome and regulated by the same maltose regulatory protein (MalT). Note that the LacI repressor protein binds only to the lac operon, which is only located at one position on the chromosome, while the Mal repressor protein binds to multiple operons (the mal regulon).
Regulons are known for operons under negative control or repression as well. For example, the arginine biosynthetic enzymes are encoded by the arginine regulon, whose operons are all under the control of the arginine repressor protein (only one of the arginine operons was shown in Figure 7.5). In regulon control, a specific DNA-binding protein binds only at those operons it controls regardless of whether it is functioning as an activator or repressor; other operons are not affected.
Many genes and regulons in E. coli have promoters under positive control and many have promoters under negative control. In addition, many operons have promoters with multiple types of control and some have more than one promoter, each with its own control system! Thus, the simple picture outlined above does not hold for all operons. Multiple control features are common in the operons and regulons of virtually all Bacteria and Archaea, and thus their overall regulation may require a network of interactions.
We now consider transcriptional control in Archaea, which has elements that both resemble and differ from those in Bacteria.
Check Your Understanding
Compare and contrast the activities of an activator protein and a repressor protein.
7.4 Transcription Controls in *Archaea*
As we have seen, there are two alternative approaches to regulating the activity of RNA polymerase. One strategy, common in Bacteria, is to use DNA-binding proteins that either block RNA polymerase activity (repressor proteins) or stimulate RNA polymerase activity (activator proteins). An alternative strategy, common in eukaryotes, is to coordinate the binding of several proteins on the DNA to interact with RNA polymerase. Because these proteins affect the rate at which a gene is transcribed, they are also considered transcription factors (Section 7.2). Given the greater overall similarity between the mechanism of transcription in Archaea and Eukarya (Section 6.6), it is perhaps surprising that the regulation of transcription in Archaea more closely resembles that of Bacteria.
Few repressor or activator proteins from Archaea have been characterized in the detail of the arg, lac, or mal operons of Escherichia coli (Figures 7.5, 7.6, 7.9, and 7.10), but it is clear that Archaea have both positive and negative control systems. Archaeal repressor proteins either block the binding of RNA polymerase itself or block the binding of TBP (TATA-binding protein) and TFB (transcription factor B), proteins that are required for RNA polymerase to bind to the promoter in Archaea (Section 6.6). At least some archaeal activator proteins function in just the opposite way, by recruiting TBP to the promoter, thereby facilitating transcription.
Control of Nitrogen Assimilation in *Archaea*
A well-studied example of an archaeal repressor is the NrpR protein from the methanogen Methanococcus maripaludis (Section 17.2). NrpR represses genes encoding nitrogen assimilation functions (Figure 7.11), such as those for nitrogen fixation (Section 3.12) and synthesis of the amino acid glutamine. When organic nitrogen is plentiful in the M. maripaludis cell, NrpR represses nitrogen assimilation genes. However, if the level of organic nitrogen becomes limiting, the compound α-ketoglutarate accumulates to high levels. This occurs because α-ketoglutarate, a citric acid cycle intermediate, is also a major acceptor of ammonia during nitrogen assimilation (Section 3.14).
Figure 7.11 Repression of genes for nitrogen metabolism in ***Archaea***.

The NrpR protein of Methanococcus maripaludis functions as a repressor. It blocks the binding of the TFB and TBP proteins, which are required for promoter recognition, to the BRE (B recognition element) site and TATA box, respectively. If there is a shortage of ammonia, α-ketoglutarate is not converted to glutamate. The α-ketoglutarate accumulates and binds to NrpR, releasing it from the DNA and allowing TBP and TFB to bind. The latter promotes RNA polymerase binding and transcription of the operon.
When levels of α-ketoglutarate rise, this signals the cell that ammonia is limiting and that additional pathways need to be activated for obtaining ammonia, such as nitrogen fixation or the activity of glutamine synthetase, the high-affinity nitrogen assimilation enzyme. Elevated levels of α-ketoglutarate function as an inducer by binding to the NrpR protein. In this state, NrpR loses its affinity for the promoter regions of its target genes and no longer blocks transcription from promoters. In this respect, the NrpR protein resembles the LacI repressor and similar-functioning proteins of Bacteria (Section 7.3 and Figure 7.6).
Dual-Acting Transcriptional Regulators in *Archaea*
Some archaeal regulators possess dual functionality and can function as both a repressor and an activator. For example, the TrmB family of transcriptional regulators that primarily regulate sugar metabolism is widespread in Archaea, with more than 250 proteins identified. The actual activity of the allosteric regulator depends on its DNA binding site. In Pyrococcus furiosus, a hyperthermophile that does not possess a glucose transporter but can perform glycolysis, TrmBL1 simultaneously functions as a repressor of genes for other sugar transport systems present in this organism and as an activator of genes for glucose synthesis (gluconeogenesis, Section 3.13).
As a repressor, TrmBL1 binds to a specific DNA sequence downstream of the BRE (B recognition element)/TATA binding sites of maltodextrin and maltose/trehalose uptake genes (**Figure 7.12*a***). This binding blocks the binding of RNA polymerase to these sites and thus prevents gene expression. If cellular conditions shift and the inducer sugars maltose, maltotriose, or fructose are present, they bind to TrmBL1, resulting in an allosteric change in the protein such that it can no longer bind DNA. Without TrmBL1 bound to the DNA, RNA polymerase is able to bind to the promoter region and derepression of sugar transporter genes occurs (Figure 7.12a).
Figure 7.12 Dual functionality of the ***Pyrococcus furiosus*** TrmBL1 regulator.

Pyrococcus furiosus is a hyperthermophilic species of the Euryarchaeota with a growth temperature optimum of 100 °C (Section 17.4). (a) The TrmBL1 protein functions as a repressor of genes for sugar uptake. It binds to DNA downstream of the TATA box, blocking the binding of RNA polymerase. Binding of maltose (or another inducer) to TrmBL1 results in release of the regulator from the DNA. Now RNA polymerase can bind and transcribe sugar uptake genes. (b) The TrmBL1 protein functions as an activator of genes for glucose synthesis. It binds to a sequence upstream of the promoter region and recruits the transcription initiation complex. This in turn results in transcription of glucose synthesis genes. The presence of maltose results in release of TrmBL1 from the DNA. Without TrmBL1 bound to the DNA, TBP and TFB are not recruited and glucose synthesis genes are not transcribed. The DNA polymerase from P. furious is a valuable tool in biotechnology (Section 12.1).
In other metabolic situations, TrmBL1 can function as an activator protein by binding to a separate and distinct DNA sequence associated with genes for glucose biosynthesis. This regulatory region differs from the one that TrmBL1 binds to as a repressor as it is located upstream of the BRE/TATA binding sites (Figure 7.12b). Binding of TrmBL1 to this DNA sequence helps recruit the archaeal transcription initiation complex (TBP, TFB, and RNA polymerase, Section 6.6), thus activating transcription. However, in the same manner as its role as a repressor, TrmBL1 as an allosteric activator will not bind to DNA if the effector molecules maltose, maltotriose, or fructose are present. Without TrmBL1 binding upstream of the BRE/TATA sequences and activating transcription, RNA polymerase and the other needed initiation factors cannot be recruited for transcription of genes necessary for glucose synthesis. This dual-functioning control conserves energy by preventing expression of the gluconeogenesis pathway when other sugars that can be converted into glucose are present in the cell.
The SurR protein of P. furiosus is another example of a regulatory protein that functions as both an activator and a repressor, depending on the location of its binding site within the promoter region. SurR controls a catabolic shift of P. furiosus from fermentation (which produces H2) to anaerobic respiration with sulfur (S0) as electron acceptor, forming H2S (Section 3.10 and Section 14.12). When S0 is absent, SurR binds to regulatory regions that activate transcription of genes that encode the enzyme hydrogenase such that P. furiosus can grow by fermentation. At the same time, SurR functions as a repressor to prevent transcription of genes encoding proteins that participate in sulfur metabolism. However, when S0 is present, SurR no longer binds to DNA. Interestingly, this inability of SurR to bind is not the result of S0 binding to SurR as an effector molecule but to the oxidation of cysteine residues within the DNA-binding domain of this regulatory protein that triggers its release from the DNA. This event both activates expression of genes that participate in S0 metabolism and represses the synthesis of hydrogenase, needed for fermentative metabolism.
Many regulatory mechanisms are triggered by environmental stimuli, and we move on to explore some of the most common ones in Part II of the chapter.
Check Your Understanding
What is the major difference between transcriptional regulation in Archaea and eukaryotes?
How do transcriptional activators in Archaea often differ in mechanism from those in Bacteria?
Explain how the Pyrococcus furiosus TrmBL1 transcription regulator is able to act as both an activator and a repressor.
II Sensing and Signal Transduction
Microbial genomes encode an assortment of multilevel regulatory mechanisms that monitor the environment for changes signaling better (or worse) growth conditions and then trigger a response that best favors survival of the cell.
Prokaryotic cells regulate cell metabolism in response to many different environmental fluctuations, including changes in temperature, pH, oxygen or nutrient availability, and even to changes in the number of other cells present. To do this, cells receive signals from the environment and transmit them to the specific target to be regulated. Some of these signals are small molecules that enter the cell and function as effectors. However, in many cases the external signal is not transmitted directly to the regulatory protein but instead is detected by a cell surface sensing system that transmits the signal to the rest of the regulatory machinery, a process called signal transduction.
7.5 Two-Component Regulatory Systems
Because most signal transduction systems contain two parts, they are called two-component regulatory systems. Characteristically, such systems consist of a specific sensor kinase protein, usually located in the cytoplasmic membrane, and a response regulator protein, present in the cytoplasm.
A kinase is an enzyme that phosphorylates compounds, typically using phosphate from ATP. Sensor kinases detect a signal from the environment and phosphorylate themselves (a process called autophosphorylation) at a specific histidine residue on the protein (Figure 7.13). Sensor kinases thus belong to the class of enzymes called histidine kinases. The phosphate is then transferred from the sensor to another protein inside the cell, the response regulator. The latter is typically a DNA-binding protein that regulates transcription in either a positive or a negative fashion (Section 7.3). In the example shown in Figure 7.13, regulation is negative; the phosphorylated response regulator functions as a repressor that binds DNA, thereby blocking transcription. Once dephosphorylated, the response regulator is released and transcription is permitted.
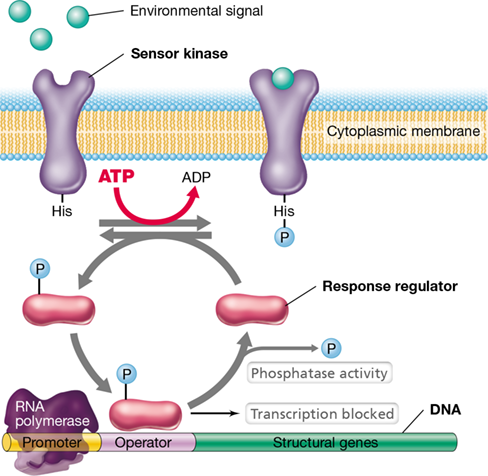
Figure 7.13 The control of gene expression by a two-component regulatory system.

One component is a sensor kinase in the cytoplasmic membrane that phosphorylates itself in response to an environmental signal. The phosphoryl group is then transferred to the second component, a response regulator. The phosphorylated form of the response regulator then binds to DNA. In the system shown here, the phosphorylated response regulator is a repressor protein. The phosphatase activity of the response regulator slowly releases the phosphate from the response regulator and resets the system.
For a balanced regulatory system to work properly, it must have a feedback loop, that is, a way to complete the regulatory circuit and terminate the response. This resets the system for another cycle. This feedback loop employs a phosphatase, an enzyme that removes the phosphate from the response regulator at a constant rate. The response regulator itself often catalyzes this reaction, although in some cases separate proteins are needed (Figure 7.13). Phosphatase activity is typically slower than kinase (phosphorylation) activity. However, if phosphorylation ceases due to reduced sensor kinase activity, phosphatase activity eventually returns the response regulator to the fully nonphosphorylated state, and the system is reset.
Examples of Two-Component Regulatory Systems
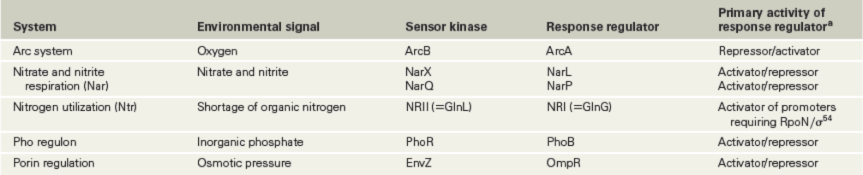
Two-component systems regulate a large number of genes in many different Bacteria, and some key examples include those that respond to phosphate limitation, nitrogen limitation, and osmotic pressure; we focus on the latter two systems here.
In Escherichia coli almost 50 different two-component systems are present, and several are listed in Table 7.1. In one example, the osmolarity (dissolved solute concentration) of the environment controls the relative levels of the proteins OmpC and OmpF in the E. coli outer membrane. OmpC and OmpF are porins, proteins that allow metabolites to cross the outer membrane of gram-negative bacteria (Section 2.4). If osmotic pressure is low, the synthesis of OmpF, a porin with a larger pore, increases; if osmotic pressure is high, OmpC, a porin with a smaller pore, is made in larger amounts.
Table 7.1 Examples of two-component systems that regulate transcription in *Escherichia coli*

aNote that many response regulator proteins act as both activators and repressors depending on the genes being regulated. Although ArcA can function as either an activator or a repressor, it functions as a repressor on most operons that it regulates.
EnvZ, a sensor histidine kinase in the E. coli cytoplasmic membrane, detects changes in osmotic pressure. When a shift occurs, EnvZ is autophosphorylated and transfers its phosphate group to OmpR, the response regulator of this system (**Figure 7.14*a***). Under conditions of low osmotic pressure, phosphorylated OmpR (OmpR-P) activates transcription of the ompF gene. Conversely, when osmotic pressure is high, OmpR-P represses transcription of ompF and simultaneously activates transcription of ompC (Figure 7.14a). The expression of ompF is also regulated by an additional control mechanism—regulatory RNA—and we consider this additional level of regulation in Section 7.12.
Figure 7.14 Two examples of well-studied two-component systems in ***Escherichia coli***.

(a) Regulation of synthesis of outer membrane proteins. The cytoplasmic membrane histidine kinase EnvZ autophosphorylates itself under osmotic pressure changes and then activates the transcriptional regulator OmpR by phosphorylation. OmpR-P binds upstream of the ompF gene and activates transcription under low osmotic pressure; conversely, it represses transcription of ompF under high osmotic pressure. OmpR-P only activates transcription of the ompC gene under conditions of high osmolarity. (b) Regulation of ammonia assimilation by the Ntr system. NRII is the sensor kinase and NRI is the response regulator. Genes encoding nitrogen assimilation proteins are only transcribed when cellular levels of ammonia are low.
Some signal transduction systems have a secondary regulatory element that is not RNA-based, and their activities can quickly become quite complex. Such is the case with the regulation of nitrogen assimilation in Bacteria. For instance, in the Ntr regulatory system (Figure 7.14b), the response regulator is an activator protein called nitrogen regulator I (NRI). In the phosphorylated state, NRI-P oligomerizes and binds to an activator-binding site over 100 base pairs upstream of the promoter region for nitrogen assimilation genes (Figure 7.14b). This binding causes the DNA to loop around such that NRI-P can make contact with RNA polymerase and the alternative sigma factor σ54 (RpoN) (Section 6.5); this activates transcription of genes that encode key nitrogen assimilation enzymes (Figure 7.7b). Phosphorylation of NRI is controlled by the sensor kinase of the Ntr system called nitrogen regulator II (NRII), which senses ammonia levels in the cell. However, NRII can switch from kinase activity to phosphatase activity depending on the nitrogen status of the cell (Figure 7.14b).
The dual enzymatic activity of NRII is controlled by the state of another protein called PII, a signal-transducing protein we will visit later when we consider regulation of an important ammonia assimilation protein, glutamine synthetase (Section 7.16). Under nitrogen starvation conditions, a nucleotide called uridine monophosphate (UMP) is added to PII, forming PII–UMP (Section 7.16). If PII–UMP is present in the cell, it stimulates the kinase activity of NRII. Thus the presence of PII-UMP leads to NRI-P and activation of genes encoding enzymes of nitrogen assimilation (Figure 7.14b). Conversely, if nitrogen is present in excess in the cell, UMP is removed from PII. PII in its unmodified form promotes the phosphatase activity of NRII, which results in unphosphorylated NRI. Without phosphorylation of NRI, transcriptional activation of nitrogen assimilation genes does not occur (Figure 7.14b). This type of regulation in which a hierarchy of control systems are deployed in a cascading fashion is common for systems of central importance to cellular metabolism.
Although rare in Archaea, two-component systems closely related to those in Bacteria are also present in microbial eukaryotes, such as the yeast Saccharomyces cerevisiae, and even in plants. However, most eukaryotic signal transduction pathways are distinct systems that phosphorylate serine, threonine, or tyrosine residues of proteins and are unrelated to those of the bacterial two-component systems that phosphorylate histidine residues (Figures 7.13 and 7.14a).
Check Your Understanding
What are kinases and what is their role in two-component regulatory systems?
What are phosphatases and what is their role in two-component regulatory systems?
7.6 Regulation of Chemotaxis
We have previously seen that some motile cells of Bacteria and Archaea respond to challenges such as nutrient limitation and toxin accumulation by moving toward attractants and away from repellents, a behavior called chemotaxis (Section 2.11). We noted there that prokaryotic cells are too small to sense spatial gradients of a chemical, but they can respond to temporal gradients. That is, they can sense the change in concentration of a chemical over time rather than the absolute concentration of the chemical stimulus. Chemotactic activity of the opportunistic pathogen Pseudomonas aeruginosa scavenging nutrients from damaged epithelial cells is shown visually in Figure 7.15.
Figure 7.15 Chemotaxis of ***Pseudomonas aeruginosa*** cells toward nutrients from damaged epithelial cells.

A time series of fluorescence micrographs illustrating epithelial cells (blue: alive; red: dead) and P. aeruginosa (green) pre– and post–epithelial cell wounding. (a) Before damage of the epithelial layer by scratching. (b) Two minutes after scratching the epithelial monolayer, very few bacterial cells are present. (c) Five minutes after wound induction, P. aeruginosa cells migrate to the damaged epithelial cells to devour nutrients released from the cells. (d) After 30 minutes, some of the epithelial cells die (red) and the bacterial hunters move on to scavenge more nutrients from living cells. Images adapted from Schwarzer, C., Fisher, H., and Machen, T.E. 2016. PLoS ONE 11(3): e0150109.
Chemotaxis has been well studied in Bacteria, which use a modified two-component regulatory system (Section 7.5) to sense temporal changes in attractants or repellents and process this information to regulate the direction of flagellar rotation. This differs from what was considered in the previous section in that the two-component system is used to regulate the activity of preexisting proteins (the flagellum protein complex) rather than to control the transcription of genes encoding flagellar proteins.
Response to Signal
The mechanism of chemotaxis depends upon a signal cascade of multiple proteins. Sensory proteins sense attractants or repellents and interact with cytoplasmic sensor kinases. These sensory proteins are called methyl-accepting chemotaxis proteins (MCPs). MCPs allow the cell to monitor the concentration of various substances over time. Thousands of MCPs cluster to form large hexagonal arrays of proteins known as chemoreceptors. Depending on the microbe, these chemoreceptors can be located in the cytoplasmic membrane and/or cytoplasm. Vibrio species, which are often associated with coastal marine waters, contain both transmembrane and cytoplasmic chemoreceptors for sensing (Figure 7.16), whereas Escherichia coli only contains transmembrane chemoreceptors.
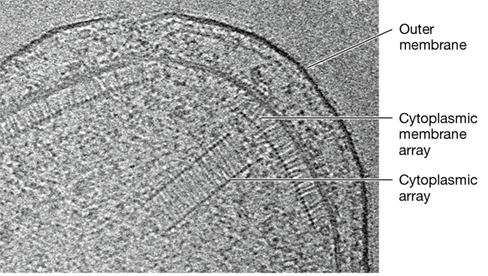
Figure 7.16 ***Vibrio*** chemoreceptor arrays.

Electron tomograph of a cell of Vibrio cholerae showing membrane-bound and cytoplasmic chemoreceptor arrays.
E. coli possesses four transmembrane chemoreceptors, each composed of five different MCPs that are specific for certain compounds. For example, the Tar MCP of E. coli senses the attractants aspartate and maltose and the repellents cobalt and nickel. MCPs bind attractants or repellents directly or in some cases indirectly through interactions with periplasmic binding proteins (Sections 2.2 and 2.4), which allows for a wider range of signals to be sensed.
MCP binding of an attractant or repellent triggers interactions with the cytoplasmic proteins CheA and CheW (Figure 7.17). CheA is the sensor kinase (Section 7.5) for chemotaxis. When MCPs bind to an attractant or release a repellent, CheW is inactive and phosphorylation of CheA is inhibited (Figure 7.17a). Thus, an increase in attractant decreases the rate of CheA autophosphorylation and the response regulator CheY remains unphosphorylated. When an MCP binds a repellent or releases an attractant, it changes conformation and, with help from CheW, leads to the autophosphorylation of CheA to form CheA-P (Figure 7.17b). Thus, an increase in repellent concentration increases the rate of CheA autophosphorylation. CheA-P then transfers the phosphate to CheY (forming CheY-P); this is the response regulator (Section 7.5) that controls flagellar rotation. As described in the next subsection, CheY-P leads to clockwise flagellar rotation. CheA-P can also transfer the phosphate to a different protein, CheB, which plays a role in adaptation (Figure 7.17), to be detailed shortly.
Figure 7.17 Mechanism of chemotaxis in ***Escherichia coli***.

Methyl-accepting chemotaxis proteins (MCPs) within chemoreceptor arrays form a complex with the sensor kinase CheA and the coupling protein CheW. (a) Binding of an attractant to MCP (or an empty MCP) inhibits CheA kinase activity. Without phosphorylation of CheY, the flagellar switch continues in a counterclockwise (CCW) rotation and cell running toward the attractant occurs. (b) Binding of a repellent to MCP stimulates CheA, which can then phosphorylate the response regulators CheB and CheY. Binding of phosphorylated CheY (CheY-P) to the flagellar motor switch results in clockwise (CW) rotation and cell tumbling. CheZ is responsible for dephosphorylating CheY-P so that the response can continue or cease depending on the activity of CheA. CheR continually adds methyl groups to the MCP, while CheB-P (but not CheB) removes them. The degree of methylation of the MCPs controls their ability to respond to attractants and repellents and leads to adaptation.
Controlling Flagellar Rotation
CheY is a central protein in the regulatory system because it governs the direction of rotation of the flagellum. Recall that if rotation of the flagellum is counterclockwise, Escherichia coli will continue to move in a run (swim smoothly), whereas if the flagellum rotates clockwise, the cell will tumble (move randomly) (Section 2.11 and Figure 2.33). When MCPs bind a repellent or release an attractant, CheY is phosphorylated. CheY-P interacts with the flagellar motor to induce clockwise flagellar rotation, which causes tumbling (Figure 7.17b). When MCPs bind an attractant or release a repellent, CheY is unphosphorylated and cannot bind to the flagellar motor. This results in the flagellum rotating counterclockwise, causing the cell to run (Figure 7.17a). Another protein, CheZ, dephosphorylates CheY, returning it to the form that allows runs instead of tumbles. Thus an increase in repellent concentration leads to an increase in the level of CheY-P and therefore tumbling. By contrast, if the cell is swimming toward attractants, the lower level of CheY-P suppresses tumbles and promotes runs.
Adaptation
Once an organism has successfully responded to a stimulus, it must stop responding and reset the sensory system to await further signals. This is known as adaptation. During adaptation of the chemotaxis system, a feedback loop resets the system. This relies on the response regulator CheB, mentioned earlier.
As their name implies, MCPs can be methylated. When MCPs are fully methylated they no longer respond to attractants but are sensitive to repellents. Conversely, when MCPs are unmethylated they respond strongly to attractants but are insensitive to repellents. Varying the methylation level thus allows adaptation to sensory signals. This is accomplished by methylation and demethylation of the MCPs. CheR is the chemotaxis protein that methylates MCPs, while phosphorylated CheB (CheB-P) is the protein that demethylates MCPs (Figure 7.17). Note that CheA-P donates a phosphate group not only to CheY as described above, but to CheB as well (Figure 7.17b).
If the level of an attractant remains high, the rate of CheA autophosphorylation is low. This leads to unphosphorylated CheY and CheB (Figure 7.17). Consequently, the cell swims smoothly in a run. Methylation of the MCPs increases during this period because CheB-P is not present to demethylate them. However, MCPs no longer respond to the attractant when they become fully methylated. Therefore, if the level of attractant remains high but constant, the cell begins to tumble. Eventually, CheB becomes phosphorylated and CheB-P demethylates the MCPs. This resets the receptors, allowing them to once again respond to further increases or decreases in the level of attractants. Put another way, the cell stops swimming if the attractant concentration is constant. It only continues to swim if ever higher levels of attractant are encountered. The course of events is just the opposite for repellents. Fully methylated MCPs respond best to an increasing gradient of repellents and send a signal for cell tumbling to begin. The cell then moves off in a random direction while MCPs are slowly demethylated. With this mechanism for adaptation, chemotaxis successfully achieves the ability to monitor small changes in the concentrations of both attractants and repellents over time.
Other Taxes
In addition to chemotaxis, several other forms of taxis are known, for example, phototaxis (movement toward light) and aerotaxis (movement toward oxygen) (Section 2.12). Interestingly, many of the cytoplasmic Che proteins that function to control flagellar activity in chemotaxis also play a role in these other taxes. For example, in phototaxis, a light sensor protein replaces the MCPs of chemotaxis, and in aerotaxis, a redox protein monitors levels of oxygen. These sensors then interact with cytoplasmic Che proteins to begin the cascade of events that direct runs or tumbles. Thus several different kinds of environmental signals converge on the same flagellar control system, and this allows the cell to economize on its regulatory systems.
Check Your Understanding
What are the primary response regulator and the primary sensor kinase for regulating chemotaxis?
How does the response of the chemotaxis system to an attractant differ from its response to a repellent?
7.7 Cell-to-Cell Signaling
Populations of Bacteria and Archaea are able to communicate with one another through the production of small extracellular molecules. Depending on the cell, these molecules can be either small peptides or nonpeptide organic molecules. Accumulation of these signaling molecules often leads to coordinated group behaviors such as biofilm formation (Section 4.9 and Section 8.10), and because accumulation is essential, the density of cells producing specific signaling molecules in a local environment ultimately controls such pathways. This regulatory phenomenon is called quorum sensing (the word “quorum” in this sense meaning “sufficient numbers”).
Mechanism of Quorum Sensing
Quorum sensing is a regulatory mechanism that assesses population density. Many bacteria use this approach to ensure that sufficient cell numbers of their own species are present before initiating activities that require a certain cell density to work effectively. For example, a pathogenic bacterium that secretes a toxin will have no effect as a single cell; production of toxin by one cell alone would merely waste the cell’s resources. However, if a sufficiently large population of cells is present, the coordinated expression of the toxin may successfully cause disease and release resources from the host that can be used to support growth of the pathogen.
Quorum sensing is widespread among gram-negative Bacteria but is also found in many gram-positive species and in some Archaea. Each species that employs quorum sensing produces a specific signal molecule called an autoinducer (Figure 7.18). This molecule typically diffuses freely across the cell envelope in either direction. Because of this, the autoinducer reaches high concentrations inside the cell only if many cells are nearby, each making the same autoinducer. In the cytoplasm, the autoinducer binds to a specific transcriptional activator protein or a sensor kinase of a two-component system (Sections 7.3, 7.4 and7.5), ultimately triggering transcription of specific genes (Figure 7.18a).
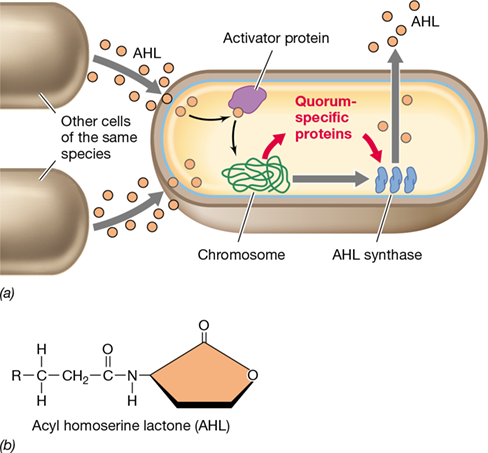
Figure 7.18 Quorum sensing.

(a) A cell capable of quorum sensing expresses acyl homoserine lactone (AHL) synthase at basal levels. This enzyme makes the cell’s specific AHL. When cells of the same species reach a certain density, the concentration of AHL rises sufficiently to bind to the activator protein, which activates transcription of quorum-specific genes. (b) General structure of an AHL. Different AHLs are variants of this parent structure. R = alkyl group (C1−C17); the carbon next to the R group is often modified to a keto group (C=O).
Mastering Microbiology
While several different classes of autoinducers exist, the first to be identified were the acyl homoserine lactones (AHLs) (Figure 7.18b). Several different AHLs, with acyl groups (functional groups containing both carbonyl and alkyl groups) of different lengths, are found in different species of gram-negative bacteria. In addition, many gram-negative Bacteria make autoinducers such as AI-2 (a cyclic furan derivative) that can be used between many different species, thus allowing more widespread gram-negative interspecies communication. Gram-positive Bacteria and some Archaea generally use certain small peptides as autoinducers.
The phenomenon of quorum sensing was discovered as the mechanism by which light emission in bioluminescent bacteria is regulated (Sections 16.4 and 23.10). Several bacterial species can emit light, including the marine bacterium Aliivibrio fischeri. Figure 7.19 shows bioluminescent colonies of A. fischeri. The light is generated by an enzyme called luciferase. The lux operons encode the proteins needed for bioluminescence and are under control of the activator protein LuxR. The lux operons are induced when the concentration of the specific A. fischeri AHL, N-3-oxohexanoyl homoserine lactone, becomes high enough. When this occurs, LuxR binds to the lux operons and activates their transcription.
Figure 7.19 Bioluminescent bacteria producing the enzyme luciferase.

Cells of the bacterium Aliivibrio fischeri were streaked on nutrient agar in a Petri dish and allowed to grow overnight. The photograph was taken in a darkened room using only the light generated by the bacteria.
Quorum sensing has also been observed in microbial eukaryotes. For example, in the yeast Saccharomyces cerevisiae, specific aromatic alcohols are produced as autoinducers and control the transition between growth of S. cerevisiae as single cells and as elongated filaments. Similar transitions are seen in other dimorphic fungi (fungi capable of two different morphologies), some of which cause disease in humans. An example is Candida, whose quorum sensing is mediated by the long-chain alcohol farnesol. As the concentration of farnesol increases, the transition from budding yeast cells to elongated hyphae is prevented.
Virulence Factors
Various genes are controlled by quorum sensing, including some in pathogenic bacteria. For example, Shiga toxin–producing Escherichia coli, such as the notorious foodborne pathogen E. coli O157:H7 (Section 33.11), produces an AHL called AI-3 that induces virulence genes. As the E. coli population increases in the intestine, bacterial cells produce AI-3 while host intestinal cells produce the stress hormones epinephrine and norepinephrine. All three of these signal molecules bind to two separate sensor kinases in the E. coli cytoplasmic membrane, resulting in the phosphorylation and activation of two transcriptional activator proteins (**Figure 7.20*a***). These proteins activate transcription of genes encoding motility functions and secretion of the toxin as well as genes encoding proteins that form lesions on the host intestinal mucosa, thus releasing nutrients that can be used by cells of the pathogen. This is a rare example of a system that senses both bacterial and eukaryotic chemical signals to regulate gene expression.
Figure 7.20 Quorum sensing regulation of virulence factors in two common bacterial pathogens.

Escherichia coli and Staphylococcus aureus can be harmless saprophytes or potent pathogens, depending on the strain. As pathogens, the production of major virulence factors is controlled by quorum sensing. (a) As the bacterial population increases, AI-3 produced by Escherichia coli and epinephrine and norepinephrine produced by the intestinal cell accumulate and bind to sensor kinases, initiating a cascade of events necessary for production of virulence factors (for example, enterotoxin). (b) Basal level transcription of the argABCD operon in Staphylococcus leads to production of ArgD, the pre-autoinducing peptide (AIP). ArgB trims ArgD into the functional AIP and exports it out of the cell. As the cell population increases, the AIP concentration increases and binds to ArgC, leading to autophosphorylation of ArgC. ArgC-P then activates the transcriptional activator ArgA by transfer of a phosphate group. ArgA-P increases transcription of the argABCD operon as well as activating the transcription of an RNA that leads to the production of virulence proteins.
The pathogenesis of the gram-positive bacterium Staphylococcus aureus (Section 31.9) requires, among many other things, the production and secretion of small extracellular peptides that damage host cells or that interfere with the host’s immune system. The genes encoding these virulence factors are under the control of a quorum-sensing system that uses a small peptide called the autoinducing peptide (AIP), encoded by the argD gene, as the autoinducer. After synthesis of ArgD (pre-AIP), the membrane-bound ArgB protein trims the peptide into its active AIP form and secretes the small peptide outside of the cell (Figure 7.20b). As the cell density of S. aureus increases, so does the concentration of AIP. ArgC is a membrane-bound sensor kinase (Section 7.5) that binds to AIP, resulting in autophosphorylation. ArgC-P then transfers its phosphate to the transcriptional activator ArgA. ArgA-P increases transcription of argABCD genes that encode the signal transduction system as well as an RNA molecule that controls production of a range of virulence proteins.
Some eukaryotes produce molecules that specifically interfere with bacterial quorum sensing. Most of these found so far have been furanone derivatives containing a halogen atom. These components mimic the AHLs or AI-2 and disrupt bacterial behavior that relies on quorum sensing. Because of this, quorum-sensing disruptors have been proposed as potential drugs to disperse bacterial biofilms (Section 4.9 and Section 8.10) and prevent the expression of virulence genes.
In the next part we explore mechanisms of controlling large gene sets scattered across the chromosome, something the cell must do to coordinate the expression of genes not linked in a single operon.
Check Your Understanding
What advantage do quorum-sensing systems confer on bacterial cells?
What properties are required for a molecule to function as an autoinducer?
How do the autoinducers used in quorum sensing by gram-negative bacteria differ from those used by gram-positive bacteria?
III Global Control
Some microbial regulatory systems control the expression of several sets of genes scattered across the genome in order to activate or repress the synthesis of groups of functionally related proteins in a single control event.
An organism often needs to regulate many unrelated genes simultaneously in response to a change in its environment. Regulatory mechanisms that respond to environmental signals by regulating the transcription of many different genes comprising more than one regulon are called global control systems. There are several global control systems in Escherichia coli (and probably in all Bacteria and Archaea), and some key ones are listed in Table 7.2. Global control networks may include activators, repressors, signal molecules, two-component regulatory systems, regulatory RNA (Section 7.12), and alternative sigma (σ) factors (Section 6.5) as components of the overall control system.
Table 7.2 Examples of global control systems known in *Escherichia coli*a

aFor many of the global control systems, regulation is complex. A single regulatory protein can play more than one role. For instance, the regulatory protein for aerobic respiration is a repressor for many promoters but an activator for others, whereas the regulatory protein for anaerobic respiration is an activator protein for many promoters but a repressor for others. Regulation can also be indirect or require more than one regulatory protein. Many genes are regulated by more than one global system.
7.8 The *lac* Operon
Both the lactose operon and the maltose regulon respond to global controls in addition to their own controls discussed in Section 7.3. We begin our consideration of global regulation by revisiting the lac operon and considering how cells respond when more than one sugar is available.
Catabolite Repression
In its intestinal habitat, Escherichia coli likely encounters several different utilizable carbon sources, and for this reason, the organism is genetically equipped to catabolize many different sugars. When given several sugars, including glucose, do cells of E. coli use them simultaneously or one at a time? The answer is that glucose is always used first. It would be wasteful to induce enzymes for using other sugars when glucose is available because E. coli grows faster on glucose than on any other carbon source. Catabolite repression is a mechanism of global control that controls the use of carbon sources if more than one is present.
When cells of E. coli are grown in a medium that contains glucose, the synthesis of enzymes needed for the breakdown of other carbon sources (such as lactose or maltose) is repressed, even if those other carbon sources are present. Thus, the presence of a favored carbon source represses the induction of pathways that catabolize other carbon sources. Catabolite repression is sometimes called the “glucose effect” because glucose was the first substance shown to cause this response. But catabolite repression is not always linked to glucose; the key point is that the favored substrate is a better carbon and energy source than other available carbon and energy sources.
Why is catabolite repression called global control? In E. coli and other organisms for which glucose is the best energy source, catabolite repression prevents expression of most other catabolic operons as long as glucose is present. Dozens of catabolic operons are affected, including those for lactose (Figure 7.6), maltose (Figure 7.9), a host of other sugars, and most other commonly used carbon and energy sources for E. coli. In addition, genes for the synthesis of flagella are controlled by catabolite repression because if bacteria have a good carbon source available, there is no need to swim around in search of nutrients.
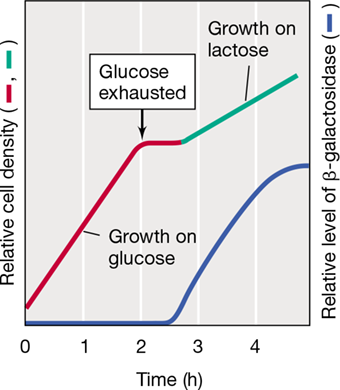
One consequence of catabolite repression is that it may lead to two exponential growth phases, a phenomenon called diauxic growth. If two usable energy sources are available, the cells first consume the better energy source. Growth stops when the better source is depleted, but following a lag period, it resumes on the other energy source. Diauxic growth is illustrated in Figure 7.21 for a culture of E. coli grown on a mixture of glucose and lactose. The cells grow more rapidly on glucose than on lactose. Although glucose and lactose are both excellent energy sources for E. coli, glucose is superior and supports a higher growth rate.
Figure 7.21 Diauxic growth of ***Escherichia coli*** on a mixture of glucose and lactose.

The presence of glucose represses the synthesis of β-galactosidase, the enzyme that cleaves lactose into glucose and galactose. After glucose is depleted, there is a lag during which β-galactosidase is synthesized. Growth then resumes on lactose but at a slower rate, as indicated by the green line.
The proteins of the lac operon, including the enzyme β-galactosidase, are required for using lactose and are induced in its presence (Figure 7.6). But the synthesis of these proteins is subject to catabolite repression, and as long as glucose is present, the lac operon is not expressed and lactose is not used. However, when glucose is depleted, catabolite repression is abolished and the lac operon is expressed; shortly thereafter, cells begin to grow on lactose. How are all of these events controlled?
Cyclic AMP and Cyclic AMP Receptor Protein
Despite its name, catabolite repression relies on an activator protein and is actually a form of activation (Section 7.3). The activator protein is called the cyclic AMP receptor protein (CRP). A gene that encodes a catabolite-repressible enzyme is expressed only if CRP binds to DNA in the promoter region. This allows RNA polymerase to bind to the promoter. CRP is an allosteric protein and binds to DNA only if it has first bound a small molecule called cyclic adenosine monophosphate (cyclic AMP or cAMP), a derivative of the nucleotide adenosine present in RNA (Figure 7.22). Like many DNA-binding proteins (Section 7.1), CRP binds to DNA as a dimer.
Figure 7.22 Cyclic AMP.

Cyclic adenosine monophosphate (cyclic AMP) is made from ATP by the enzyme adenylate cyclase.
Cyclic AMP is a key molecule in many metabolic control systems, both in prokaryotic cells and eukaryotes. Because it is derived from a nucleic acid precursor, it is a regulatory nucleotide. Other regulatory nucleotides include cyclic guanosine monophosphate (cyclic GMP; important mostly in eukaryotes), cyclic di-GMP (important in biofilm formation; Section 4.9 and Section 8.10), and guanosine tetraphosphate (ppGpp, important in the stringent response, Section 7.9). Cyclic AMP is synthesized from ATP by an enzyme called adenylate cyclase (Figure 7.22). However, glucose inhibits the synthesis of cyclic AMP and also stimulates cyclic AMP transport out of the cell. When glucose enters the cell, the cyclic AMP level is lowered, CRP cannot bind DNA, and RNA polymerase fails to bind to the promoters of operons subject to catabolite repression. Thus, catabolite repression is an indirect result of the presence of a better energy source (glucose); the direct cause of catabolite repression is a low level of cyclic AMP.
Let us return to the lac operon and include catabolite repression. The entire regulatory region of the lac operon is diagrammed in Figure 7.23. For lac genes to be transcribed, two requirements must be met: (1) The level of cyclic AMP must be high enough for the CRP protein to bind to the CRP-binding site (positive control), and (2) lactose or another suitable inducer must be present so that the lactose repressor (LacI protein) does not block transcription by binding to the operator (negative control). If these two conditions are met, the cell is signaled that glucose is absent and lactose is present; then and only then does transcription of the lac operon begin.
Figure 7.23 Overall regulation of the ***lac*** system.

The lac operon consists of lacZ, encoding β-galactosidase, lacY, encoding lactose permease, and lacA, encoding lactose acetylase. The LacI repressor protein is encoded by a separate gene, lacI. LacI binds to the operator (O) unless the inducer is present. The cyclic AMP receptor protein (CRP) binds to the CRP activator site (C) when activated by cyclic AMP and recruits RNA polymerase to bind to the promoter (P). For the lac operon to be transcribed by RNA polymerase, the LacI repressor must be absent (that is, inducer must be present) and cyclic AMP levels must be high (due to the absence of glucose), allowing CRP to bind.
Mastering Microbiology
Art Activity: Figure 7.23 Overall regulation of the lac system
We now consider another form of global control distinct from catabolite repression but where regulatory nucleotides play a central role.
Check Your Understanding
Explain how catabolite repression depends on an activator protein.
What role does cyclic AMP play in glucose regulation?
Explain how the lac operon is both positively and negatively controlled.
7.9 Stringent and General Stress Responses
In nature microorganisms must often survive nutrient-limited conditions and exposure to environmental stressors such as extreme pH, oxidative stress, and antibiotic exposure. How do cells cope and adapt to such conditions? While some gram-positive bacteria undergo sporulation to withstand such harsh conditions (Section 2.8), many Bacteria instead activate a variety of stress survival pathways—including both specific and general—that improve their chances of survival.
Mechanism of the Stringent Response
The stringent response is a widely distributed regulatory mechanism used by bacteria to survive nutrient deprivation, environmental stresses, and antibiotic exposure. Triggering of the stringent response ultimately leads to a shutdown of macromolecule synthesis and the activation of pathways to improve the cell’s ability to survive and compete in nature.
Nutrient levels for microbes in nature can change significantly and rapidly. Such changing conditions can easily be simulated in the laboratory, and much work has been done on the regulation of gene expression following a “shift down” or “shift up” in nutrient status. These include, in particular, the regulatory events triggered by starvation for amino acids or energy, experiments that led to the discovery of the stringent response.
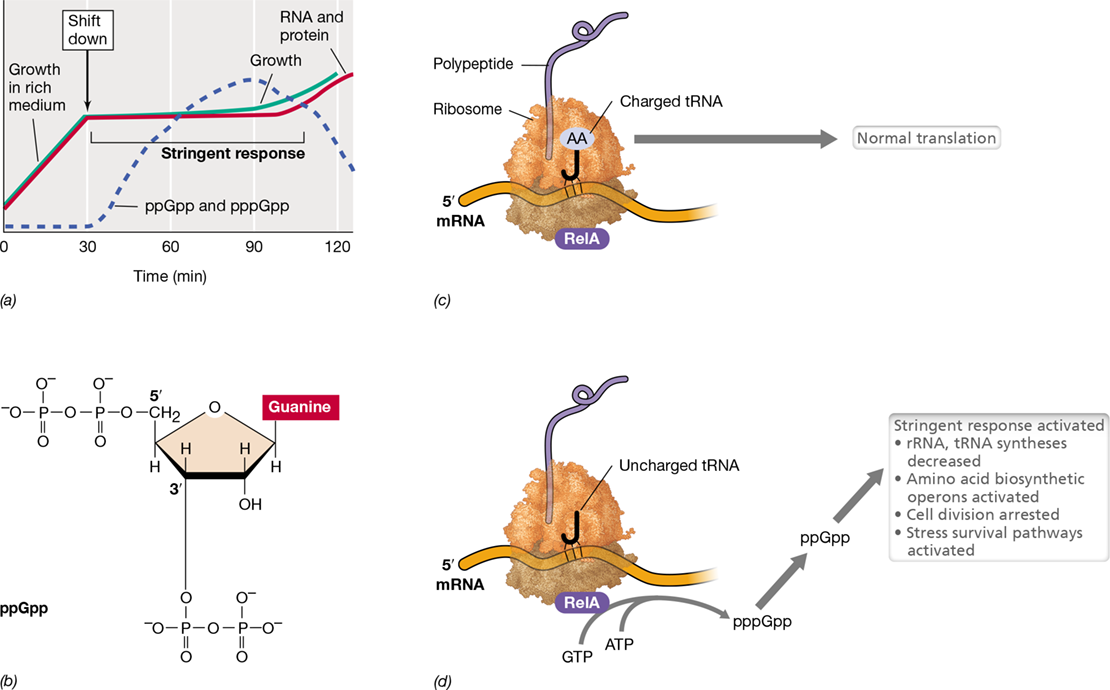
As a result of a shift down from amino acid excess to limitation, as occurs when a culture is transferred from a rich complex medium to a defined medium with a single carbon source (Table 4.2), the synthesis of rRNA and tRNA ceases almost immediately, and no new ribosomes are produced (**Figure 7.24*a***). Protein and DNA synthesis are also curtailed, but the biosynthesis of new amino acids is activated. Following such a shift, new proteins must be made to synthesize the amino acids no longer available in the environment, and these proteins are made by existing ribosomes. After a while, rRNA synthesis (and hence, the production of new ribosomes) begins again but at a new rate commensurate with the cell’s reduced growth rate (Figure 7.24a). This course of events is called the stringent response (or stringent control) and, like catabolite repression (Section 7.8), is another example of global control.
Figure 7.24 The stringent response in ***Escherichia coli***.

(a) Upon nutrient downshift, rRNA, tRNA, and protein syntheses temporarily cease. Sometime later, growth resumes at a decreased rate. (b) Structure of guanosine tetraphosphate (ppGpp), a trigger of the stringent response. (c) Normal translation, which requires charged tRNAs. (d) Synthesis of ppGpp. When cells are starved for amino acids, an uncharged tRNA can bind to the ribosome, which stops ribosome activity. This event triggers the RelA protein to synthesize a mixture of pppGpp and ppGpp.
The stringent response is triggered by a mixture of two regulatory nucleotides, guanosine tetraphosphate (ppGpp, Figure 7.24b) and guanosine pentaphosphate (pppGpp) [collectively, these are referred to as (p)ppGpp)]. In Escherichia coli, these nucleotides, which are also called alarmones, rapidly accumulate during stress or a shift down from amino acid excess to amino acid starvation. Alarmones are synthesized by a specific protein, called RelA, using ATP as a phosphate donor (Figure 7.24c, d). RelA adds two phosphate groups from ATP to GTP or GDP, thus producing pppGpp or ppGpp, respectively. RelA is associated with the 50S subunit of the ribosome and is activated by a signal from the ribosome during amino acid limitation. When the growth of the cell is limited by a shortage of amino acids, the pool of uncharged tRNAs increases relative to charged tRNAs (Section 6.8). Eventually, an uncharged tRNA is inserted into the ribosome instead of a charged tRNA during protein synthesis. When this happens, the ribosome stalls, and this leads to (p)ppGpp synthesis by RelA (Figure 7.24d). The protein Gpp converts pppGpp to ppGpp so that ppGpp is the major overall product.
The alarmones ppGpp and pppGpp have global control effects. They strongly inhibit rRNA and tRNA synthesis by binding to RNA polymerase and preventing initiation of transcription of genes for these RNAs in gram-negative bacteria. In gram-positive bacteria, the same alarmones have been shown to interfere with initiating ribonucleotides for transcription. In both gram-negative and gram-positive Bacteria, the alarmones activate both the stress response pathways and the biosynthetic operons for certain amino acids. In E. coli, the stringent response also inhibits the initiation of new rounds of DNA synthesis and cell division and slows down the synthesis of cell envelope components, such as membrane lipids.
In addition to RelA, a second protein called SpoT helps trigger the stringent response. SpoT can either make (p)ppGpp or degrade it. Under most conditions, SpoT is responsible for degrading (p)ppGpp; however, SpoT synthesizes (p)ppGpp in response to certain stresses or when nutrient (and thus energy) deprivation is detected. Thus the stringent response results not only from the absence of precursors for protein synthesis, but also from the lack of energy for biosynthesis.
The Stringent Response and Microbial Ecology
Because the stringent response is a global mechanism that balances the metabolic state of the cell, the environment or habitat of the bacterium ultimately determines the response cascade. When cells of E. coli are voided in the feces and face a switch from the nutrient-rich intestine to an open water system, the reduction in nutrients triggers ppGpp synthesis to initiate the stringent response (**Figure 7.25*a***).
Figure 7.25 Environmental control of stringent response signaling in ***Escherichia coli*** and ***Caulobacter crecentus*** (both gram-negative bacteria), and ***Mycobacterium tuberculosis*** (a gram-positive bacterium).

(a) Escherichia coli. When E. coli cells move from the nutrient-rich intestine to an open water source, they experience nutrient limitation. As ppGpp production increases, cell division is inhibited. (b) Caulobacter crescentus. C. crescentus naturally inhabits freshwater systems that are limited in nutrients. If cells encounter severe nutrient limitation, the stringent response is induced and ppGpp production increases. This increase leads to the cell morphology changing from stalked cells to swarmer cells that can swim to find more nutrients. (c) Mycobacterium tuberculosis. When M. tuberculosis cells enter a host, the bacterial cells are exposed to the host’s immune response. This leads to production of ppGpp, which converts some cells to relatively dormant persister cells. These persister cells are resistant to antibiotics and can dominate more sensitive vegetative cells, forming granulomas. Once persister cells are aerosolized and relieved of stress, they revert back to infective vegetative cells. Unlike many other bacterial pathogens that acquire antibiotic resistance from mobile genetic elements, horizontal gene transfer is rare in M. tuberculosis. Antibiotic resistance in this pathogen is primarily due to chromosomal mutations affecting the target of the antibiotic.
In contrast to E. coli, the aquatic bacterium Caulobacter naturally inhabits oligotrophic (nutrient-poor) freshwaters, and in this bacterium, the stringent response is triggered by carbon and ammonia starvation, not amino acid limitation. Caulobacter can form two types of cells, stalked cells and swarmer cells (Figure 7.25b). To increase its probability of survival, the production of ppGpp in Caulobacter leads to an increase in swarmer cell formation. Unlike stalked cells, swarmer cells are motile and can therefore swim and perhaps reach a niche containing more nutrients ( Section 8.8; Figure 7.25b).
In a disease example, the stringent response has also been implicated in the persistence of the aerobic bacterial pathogen Mycobacterium tuberculosis (tuberculosis, Section 31.4) in dormant granulomas. Within the lung of the human tuberculosis patient, the local environment is both hypoxic (low in O2) and limited for phosphate, and these conditions trigger the stringent response in cells of M. tuberculosis. Activation of the stringent response also leads to a subpopulation of M. tuberculosis cells that convert to dormant persister cells (Section 8.12); these cells are resistant to antibiotics and can survive in granulomas that form in the lung (Figure 7.25c). Once drug treatment of the infection has greatly reduced cell numbers and the environmental stresses have been relieved, the M. tuberculosis persister cells can then revert back to infective cells and trigger chronic tuberculosis, a not uncommon occurrence and one of the reasons drug therapy for tuberculosis is carried out over a long period.
The General Stress Response: The RpoS Regulon
Bacterial cells often respond to nutrient limitation and stressors by entering into the stationary phase of growth (Section 4.6). This growth switch is controlled by activating a regulon recognized by the alternative sigma factor RpoS (σS or σ38, Table 6.3), also called the stationary phase sigma factor. The RpoS regulon comprises over 400 genes including those associated with nutrient limitation, resistance to DNA damage, biofilm formation, and responses to osmotic, oxidative, and acid stresses. Thus, RpoS not only senses environmental changes but also relays signals to other regulators.
Examples of E. coli genes that are recognized by RpoS include dinB, which encodes DNA polymerase IV of the SOS DNA repair system (Section 9.4 and Figure 9.10) and catalase genes necessary for combating reactive oxygen species (Section 4.16). Because of the expansive nature of the RpoS regulon, this alternative sigma factor is itself controlled by several transcriptional, translational, and post-translational regulatory mechanisms. For example, transcription of the rpoS gene increases in response to the presence of the alarmone ppGpp, thus directly linking the RpoS regulon with the stringent response. Translation of rpoS mRNA is also positively regulated by the presence of small RNAs expressed during stress conditions (Section 7.12). Conversely, the RpoS protein is susceptible to degradation during nonstressful conditions. Overall, the general stress response highlights the global control nature of RpoS and the coordinated processes that a cell must undertake for adaptation and survival in nature.
We next consider a more specific global regulatory mechanism focused on phosphate, an essential nutrient for life (Section 4.1).
Check Your Understanding
Which Escherichia coli genes are activated and which are repressed during the stringent response, and why?
How are the alarmones ppGpp and pppGpp synthesized?
What are some other conditions that trigger the stringent response in other bacteria?
7.10 The Phosphate (Pho) Regulon
Phosphorus is not only essential for DNA, RNA, and membrane synthesis, but it is also critical for energy generation (Section 4.1) and cell signaling. In nature, phosphorus is generally in the form of inorganic phosphate (PO43−, abbreviated Pi) and is often the limiting nutrient in many environments. Because of this, Pi transformations are heavily regulated. The phosphate (Pho) regulon consists of a two-component regulatory system (Section 7.5) that responds to Pi limitation. This global control system includes not only genes encoding extracellular enzymes for extracting Pi from organic phosphates, but also genes encoding Pi transporters, enzymes for Pi storage, and other energy-requiring metabolic and biosynthesis pathways as well.
*Streptomyces* and Phosphate
Streptomyces is a genus of gram-positive Bacteria that contains many species that produce antibiotics (Section 16.12) and whose phosphate metabolism has been well studied. While antibiotic production is an energy-intensive process, the process is limited by high phosphate concentrations. In the environment, Streptomyces produce antibiotics to kill competing cells. Thus, low phosphate levels signal Streptomyces that competition for resources is keen, and this results in antibiotic production.
In Streptomyces, the two-component Pho regulatory system that senses Pi limitation consists of a membrane-bound histidine kinase sensor protein PhoR and a cytoplasmic transcriptional regulator PhoP. While the mechanism for actually sensing extracellular Pi levels is unknown, low environmental Pi levels trigger PhoR kinase activity, yielding a phosphorylated PhoP (PhoP-P) (Figure 7.26). Once phosphorylated, PhoP-P binds to conserved promoter regions called “Pho boxes” scattered across the genome. This binding signals the major sigma subunit of Streptomyces RNA polymerase to bind, and activates transcription of genes needed for Pi uptake. The same event occurs with promoters for some genes that encode products necessary for antibiotic production. By contrast, when Pi is present in excess, PhoP is dephosphorylated, and this leads to the removal of PhoP from Pho boxes and a decreased transcription of genes encoding proteins of phosphate metabolism and antibiotic synthesis.
Figure 7.26 The phosphate (Pho) regulon of ***Streptomyces***.

If inorganic phosphate (Pi) concentrations are low, the kinase PhoR is signaled to phosphorylate PhoP. PhoP-P recognizes and binds to several promoters, functioning as either an activator or a repressor. This regulation results in activating phosphate uptake and antibiotic synthesis genes and decreasing nitrogen metabolism by repressing genes regulating glutamine synthetase (glnA) and ammonium transport (amtB), respectively.
While the Pho regulon is the mechanism many Bacteria use to respond to Pi starvation, why is this regulon considered an example of global control? Besides its role as an activator of Pi uptake and antibiotic synthesis genes in Streptomyces and of Pi genes in many other Bacteria, PhoP-P also binds some promoters weakly without recruiting a sigma subunit. This binding blocks RNA polymerase and thus leads to repression of the linked gene. Somewhat surprisingly, PhoP-P appears to actually repress more genes than it activates. Genes regulated by Pi limitation and PhoP-P that are unassociated with Pi uptake in Streptomyces include those whose products are necessary for nitrogen metabolism (glutamine synthetase, glnA; ammonium transport, amtB) (Figure 7.26). The physiological rationale here is that if Pi levels are growth limiting, the cell’s production of key biosynthetic intermediates (such as amino acids) should be reduced until Pi levels increase.
Pathogenesis and the Pho Regulon
The Pho regulon can also control some aspects of pathogenesis. Pathogenic bacteria are exposed to both Pi-limiting and Pi-rich environments within a host depending on the site of infection. The genome of Shiga toxin–producing Escherichia coli, such as the notorious foodborne pathogen E. coli O157:H7 (Section 33.11), contains a Pho regulon of over 40 genes. In its normal habitat—the intestinal tract—E. coli often experiences phosphate limitation, and this condition activates the cytoplasmic-membrane-bound PhoR. This protein then transfers a phosphate group to the cytoplasmic transcriptional regulator and DNA-binding protein PhoB (the equivalent to the Streptomyces PhoP, Figure 7.26). PhoB-P triggers increased expression not only of the genes for phosphate metabolism, but also of genes encoding the acid shock protein Asr and Shiga toxin. Increased production of Asr leads to protection of periplasmic proteins from acid stress, which would be encountered by E. coli cells present in the stomach en route to the intestine. Once the pathogen is inside an intestinal cell, its Shiga toxin production is responsible for the severe diarrhea associated with the infection, an event that effectively transmits the pathogen to a new host. Thus, phosphate limitation leads to increased fitness and survivability of pathogenic strains of E. coli.
While the mechanisms remain unclear, the Pho regulon has also been implicated in regulating biofilm formation (Section 4.9 and Section 8.10), antimicrobial resistance (Section 28.7), and toxin production (Section 25.6) in other pathogenic bacteria such as Vibrio cholerae and Pseudomonas species. Thus, although the Pho regulon is the major barometer of Pi starvation, its activities impact a wide range of unrelated genes that control several distinct cellular functions.
We move on now to the last section of Part III to consider globally regulated systems found in all domains of life that respond to major temperature shifts, the phenomenon of heat shock.
Check Your Understanding
What category of regulatory system is the Pho regulon?
What types of bacterial processes are activated and repressed by phosphate limitation?
7.11 The Heat Shock Response
Most proteins are relatively stable, even to small increases in temperature. However, some proteins are less stable at elevated temperatures and tend to unfold (denature, Section 6.7). Other stress conditions besides heat can lead to protein denaturation. These include exposure to high levels of certain solvents (for example, ethanol), osmotic stress, and high doses of ultraviolet (UV) light. Because protein stability is critical for survival, cells employ a global control mechanism called the heat shock response to protect cells from protein denaturation. This response features many of the regulatory modes illustrated in Figure 7.1.
Heat Shock Proteins
Temperature and various stress conditions can generate large amounts of inactive proteins that need to be either refolded (and in the process, reactivated) or degraded to release free amino acids for the synthesis of new proteins (enzymes called proteases facilitate the latter process). Consequently, heat stress and other conditions that lead to protein unfolding trigger the synthesis of a set of proteins known as heat shock proteins. Heat shock proteins help counteract protein damage and assist the cell in recovering from stress.
In most Bacteria and Archaea there are five major classes of heat shock protein: Hsp100, Hsp90, Hsp70, Hsp60, and Hsp10. We have encountered some of these proteins before, although not by these names (Section 6.11 and Figure 6.40a). The Hsp70 protein of Escherichia coli is DnaK, which prevents aggregation of newly synthesized proteins and stabilizes unfolded proteins. Major representatives of the Hsp60 and Hsp10 families in E. coli are the proteins GroEL and GroES, respectively. These are molecular chaperones that catalyze the correct refolding of misfolded proteins. Another group of heat shock proteins (Hsp100) includes various proteases that degrade denatured or irreversibly aggregated proteins. Because heat shock proteins perform vital functions in the cell, a low level of these proteins is always present, even under optimal conditions. However, genes encoding many bacterial heat shock proteins also make up a regulon (Section 7.3) whose expression is increased by the presence of an alternative sigma factor known as RpoH.
The Alternative Sigma Factor RpoH
In many bacteria, such as E. coli, the heat shock response is controlled by the alternative RNA polymerase σ factor (Section 6.5) RpoH (σ32) (Figure 7.27). RpoH controls expression of heat shock proteins and is normally degraded within a minute or two of its synthesis. However, when cells suffer a heat shock (or other stressor that leads to protein denaturation), degradation of RpoH is inhibited and its level therefore increases. As a consequence, transcription of those operons whose promoters are recognized by RpoH increases.
Figure 7.27 Control of heat shock in ***Escherichia coli***.

(a) The mRNA that encodes the alternative sigma factor RpoH folds under normal conditions, thus blocking the ribosome-binding site (RBS) and limiting translation. The RpoH protein that is made is broken down rapidly by proteases at 32 °C, and binding of the DnaK chaperone to RpoH stimulates this degradation. (b) At 40 °C, the mRNA encoding RpoH unfolds and more RpoH protein is made. The shift in temperature also denatures some proteins and DnaK recognizes, binds, and refolds these unfolded polypeptide chains. This removes RpoH from DnaK, which slows the degradation rate of RpoH. When the level of RpoH rises, the heat shock genes are transcribed.
The rate of RpoH degradation depends on the level of free DnaK, which inactivates RpoH. In unstressed cells, the level of free DnaK is relatively high and the level of intact RpoH is correspondingly low (Figure 7.27a). However, if heat begins to unfold proteins, DnaK binds preferentially to unfolded proteins and so is no longer free to degrade RpoH (Figure 7.27b). Thus, the more denatured proteins there are, the lower the level of free DnaK and the higher the level of RpoH; the result is heat shock gene expression. When the environmental stressor has passed, for example upon a temperature downshift, RpoH is once again inactivated by DnaK and the synthesis of heat shock proteins is reduced. Thus, DnaK functions as a cellular “thermometer” for the heat shock response and its inactivation of RpoH is considered a form of post-translational regulation, which we will discuss in Section 7.16.
Not only is the activity of the RpoH protein controlled, but translation of its corresponding mRNA is also regulated. Under normal cellular conditions, the rpoH mRNA base-pairs within itself (Figure 7.27a). This folding results in a secondary structure that decreases the ribosome’s ability to bind to the ribosome-binding site (RBS) of the mRNA (Section 6.9). However, when the cell’s internal temperature increases, the mRNA encoding RpoH unfolds. This unfolding allows the ribosome easier access and thus more overall RpoH protein production under the heat shock response (Figure 7.27b).
There is also a heat shock response in Archaea, even in species that grow optimally at very high temperatures. An analog of the bacterial Hsp70 is found in many Archaea and is structurally similar to that present in gram-positive species of Bacteria. However, other types of heat shock proteins unrelated to the stress proteins of Bacteria are also present in Archaea, Moreover, a heat shock response also exists in eukaryotes. Thus, this critically important mechanism for salvaging damaged yet recoverable proteins is present in all domains of life.
Check Your Understanding
How do levels of RpoH control the heat shock response?
IV RNA-Based Regulation
**In the course of evolution, prokaryotic cells have missed out on few if any potential regulatory mechanisms. For example, controls employing RNAs (rather than proteins) to monitor the cell’s status and trigger specific gene expression are known and are widespread in Bacteria and Archaea.**
Thus far we have focused on regulatory mechanisms in which proteins sense signals or bind to DNA. In some cases, a single protein does both; in other cases, separate proteins carry out these two activities. Nonetheless, all of these mechanisms rely on regulatory proteins. However, some regulatory mechanisms employ RNA to regulate gene expression, both at the level of transcription and at the level of translation, and we consider these now.
7.12 Regulatory RNAs
RNA molecules that are not translated to give proteins are collectively known as noncoding RNA (ncRNA). This category includes the rRNAs and tRNAs that participate in protein synthesis and the RNA present in the signal recognition particle that catalyzes some types of protein secretion (Sections 6.8, 6.10, and 6.12). Noncoding RNA also includes small RNA molecules necessary for RNA processing, especially the splicing of mRNA in eukaryotes (Section 6.6). Small RNAs (sRNAs) that range from 40 to 400 nucleotides long and regulate gene expression are widely distributed in both prokaryotic cells and eukaryotes. In Escherichia coli, for example, a number of sRNA molecules regulate several aspects of cell physiology in response to environmental or cellular signals by binding to other RNAs or in some cases to other small molecules; the end result is control of gene expression.
Small RNAs (sRNAs) exert their effects by base-pairing directly to other RNA molecules, usually mRNAs, in regions of complementary sequence. This binding immediately modulates the rate of target mRNA translation because a ribosome cannot translate double-stranded RNA. Thus, sRNAs provide an additional mechanism to regulate the synthesis of a protein once its corresponding mRNA has already been made.
Mechanisms of sRNA Activity
Small RNAs alter the translation of their mRNA target by four different mechanisms (Figure 7.28). Some sRNAs base-pair to their target mRNA, changing its secondary structure to either block a previously accessible ribosome-binding site (RBS) (Section 6.9) or to open up a previously blocked RBS, allowing access for the ribosome. These two events decrease or increase expression of the protein encoded by the target mRNA, respectively. The other two mechanisms of sRNA interaction affect mRNA stability; binding of the sRNA to its target either increases or decreases degradation of the transcript by bacterial ribonucleases, thus modulating protein expression. Increased degradation of an mRNA prevents the synthesis of new protein molecules encoded by that mRNA. Alternatively, increasing the stability of mRNA will lead to higher corresponding protein levels in the cell (Figure 7.28).
Figure 7.28 Small RNA mechanisms for modulating the translation of mRNA.

(a) Binding of a ribosome to mRNA requires that the ribosome-binding site (RBS) of the mRNA be single-stranded. Binding of an sRNA to the RBS (shown in 1) can prevent translation while the binding of an sRNA to an mRNA whose RBS has secondary structure (shown in 2) can stimulate translation. (b) Ribonuclease degrades RNA. Ribonuclease binding to partially double-stranded RNA results in RNA degradation (shown in 1), while sRNA binding at the ribonuclease binding site (shown in 2) can protect the mRNA from degradation. Compare the mechanisms for controlling translation by small RNAs shown in this figure with the mechanism of controlling translation by a riboswitch shown in Figure 7.31a.
Types of Small RNA
Small RNAs are important modulators of various cellular processes including oxidative, iron limitation, and glucose–phosphate stresses as well as quorum sensing, biofilm formation, and stationary phase events. Transcription of sRNAs is often enhanced under conditions in which their target genes need to be silenced. For example, the RyhB RNA of Escherichia coli is transcribed when iron is limiting for growth. RyhB sRNA binds to several distinct target mRNAs that encode proteins needed for iron metabolism or that use iron as cofactors. Binding of RyhB sRNA blocks the RBS of the mRNA and thus inhibits translation (Figure 7.28a). The base-paired RyhB/mRNA molecules are then degraded by ribonucleases, in particular, ribonuclease E, before translation of the mRNA can occur. Small RNA control forms only part of the mechanism by which E. coli and related bacteria reduce the production of iron-requiring proteins in response to iron limitation. Other responses include transcriptional controls by repressor and activator proteins (Section 7.2) that decrease or increase, respectively, the capacity of cells to take up iron or to tap into intracellular iron reserves.
Similarly, SgrS is an sRNA in E. coli that is expressed during glucose–phosphate stress to avoid accumulation of glucose 6-phosphate. High levels of glucose 6-phosphate within the cell can block the glycolytic pathway (Section 3.6) and ultimately lead to cell death if not overcome. When SgrS is expressed, the sRNA molecule binds to the ptsG mRNA encoding a glucose transporter. This binding results in a double-stranded RNA complex that triggers degradation of ptsG mRNA by ribonuclease E, as shown in living cells in Figure 7.29. This degradation decreases the amount of ptsG mRNA that is translated, and thus lower levels of the glucose transporter protein are made. This in turn results in decreased cellular levels of glucose 6-phosphate (recall that glucose is transported into E. coli by the phosphotransferase system, which converts glucose into glucose 6-phosphate during the transport process, Section 2.2 and Figure 2.6). By reducing glucose 6-phosphate levels, the glycolytic pathway remains active.
Figure 7.29 The interaction of SgrS sRNA and ***ptsG*** mRNA during glucose– phosphate stress.

Three-dimensional super-resolution image of fluorescently labeled SgrS sRNA (red) and ptsG mRNA (green) in cells of Escherichia coli. (a) Before glucose–phosphate stress at time 0, ptsG mRNA is more abundant than SgrS sRNA. (b) After 10 min of glucose–phosphate stress, SgrS levels are higher, which leads to degradation of ptsG mRNA.
Because many sRNAs, including RyhB and SgrS, are encoded in intergenic regions and can be spatially separated from their mRNA target, they are called trans-sRNAs. As such, these sRNAs usually have limited complementarity to their target molecule and may only base-pair with a 5- to 11-nucleotide stretch. The binding of trans-sRNAs to their targets often depends on a small protein called Hfq (Figure 7.30) that binds to both mRNA and its corresponding sRNA to facilitate their interaction. Hfq forms hexameric rings with RNA-binding sites on both surfaces. Hfq and functionally similar proteins are called RNA chaperones, as they help small RNA molecules, including many sRNAs, maintain their correct structure (Figure 7.30).
Figure 7.30 The RNA chaperone Hfq holds RNAs together.

Binding of sRNA to mRNA often requires assistance from the Hfq protein. Small RNA molecules usually have several stem–loop structures. One consequence is that the complementary base sequence that recognizes the mRNA is noncontiguous.
Small RNAs do not always work by affecting mRNA. For example, replication of the ColE1 plasmid in Escherichia coli is regulated by an sRNA that primes DNA synthesis on the plasmid and its antisense partner that blocks initiation of DNA synthesis. The level of the antisense RNA determines how often replication is initiated. Some sRNAs also bind directly to proteins and modulate their activity. Thus, depending on the sRNA, these molecules can affect several different activities in the cell.
In the next section we examine a different form of RNA-based regulation from that we have considered here with the sRNAs.
Check Your Understanding
How do sRNAs alter the translation of target mRNAs?
Why do trans-sRNAs often require a chaperone protein?
How does SgrS help Escherichia coli prevent a potential metabolic disaster?
7.13 Riboswitches
RNA can carry out many roles once thought to be limited to proteins. In particular, RNA can specifically recognize and bind other molecules, including low-molecular-weight metabolites. It is important to emphasize that such binding does not require complementary base pairing (as does binding of the sRNAs described in the previous section) but results from the folding of the RNA into a specific three-dimensional structure that recognizes the target molecule, much as a protein enzyme recognizes its substrate in its active site. Catalytically active RNAs are called ribozymes. Other RNA molecules resemble repressors and activators in binding small metabolites and regulating gene expression; these are the riboswitches, our focus here.
Mechanisms of Riboswitches
Earlier in this chapter we discussed the regulation of gene expression by negative control of transcription (Section 7.3). In this process, a specific metabolite interacts with a specific repressor protein to prevent transcription of genes encoding enzymes for the biosynthetic pathway of the metabolite. In contrast, in a riboswitch there is no regulatory protein. Instead, the metabolite binds directly to a specific mRNA. This binding results in the riboswitch controlling gene expression at either the transcriptional or translational level, depending on the type of riboswitch.
Riboswitch RNAs have sequences upstream of their coding regions (5′-UTR; Figure 7.1) that can fold into specific three-dimensional structures that bind small molecules. These recognition domains are the aptamer region or “switch” in the riboswitch and exist as two alternative secondary structures, one with the small molecule bound and the other without (Figure 7.31). Alternation between the two forms of the riboswitch thus depends on the presence or absence of the small molecule, which in turn controls the secondary structure of the downstream RNA called the expression platform of the riboswitch. In one type of riboswitch, the secondary structure of the expression platform controls whether the ribosome binds to the corresponding mRNA’s ribosome-binding site (RBS) and begins translation (Figure 7.31a); in another type, it controls whether RNA polymerase continues to transcribe the mRNA (Figure 7.31b). Riboswitches exist in all three domains of life, with over 40 different types known in Bacteria. Riboswitches control not only the synthesis of enzymes in biosynthetic pathways for various vitamins, a few amino acids, some nitrogen bases of nucleic acids, and a peptidoglycan precursor, but also aspects related to biofilm formation, fluoride toxicity, and molybdenum/tungsten cofactor metabolism (Table 7.3).
Figure 7.31 Regulation by a riboswitch.

Riboswitches are located in the 5′ untranslated region (UTR) of mRNAs (Figure 7.1). The aptamer region of the riboswitch binds specifically to the metabolite. Numbers indicate regions within the riboswitch expression platform that can base-pair together, and RBS indicates the ribosome-binding site. (a) Translational regulation: Binding of a specific metabolite alters the secondary structure of the riboswitch domain, preventing translation. (b) Transcriptional regulation: Binding of a specific metabolite alters the secondary structure of the riboswitch domain, which results in the formation of a transcriptional terminator, preventing transcription of the coding region of the gene. Compare this process to that of attenuation shown in Figure 7.33a.
Table 7.3 Riboswitches in biosynthetic pathways of *Escherichia coli*

The metabolite that is bound by the riboswitch aptamer region is typically the product of a biosynthetic pathway whose constituent enzymes are encoded by the mRNAs that carry the corresponding riboswitches. For example, the thiamine riboswitch that binds the vitamin thiamine pyrophosphate lies upstream of the coding sequences for enzymes that participate in the thiamine biosynthetic pathway. When the pool of thiamine pyrophosphate is sufficient in the cell, this metabolite binds to the aptamer region of its specific riboswitch mRNA. This binding results in a secondary structure of the riboswitch that blocks the ribosome-binding site on the mRNA; this prevents translation (Figure 7.31a). If the concentration of thiamine pyrophosphate drops sufficiently low, the molecule dissociates from its riboswitch mRNA, and this unfolds the mRNA and exposes the ribosome-binding site, allowing the mRNA to bind to the ribosome and be translated.
In contrast to riboswitches that control translation, some riboswitches control transcription. This mechanism is similar to regulation by attenuation, to be discussed in Section 7.14. With transcriptional riboswitches, metabolite binding to the aptamer region results in a conformational change within the expression platform that causes RNA polymerase to prematurely terminate synthesis of the mRNA (Figure 7.31b). This occurs because metabolite binding converts the expression platform to a form that mimics a traditional transcriptional terminator (Section 6.5). The SAM (S-adenosylmethionine) riboswitch associated with sulfur metabolism in gram-positive bacteria such as Bacillus is a good example of a riboswitch that controls transcription in this way. SAM is a cofactor in the methionine biosynthesis pathway and its availability is a signal that sufficient methionine levels exist in the cell. Binding of SAM to the SAM riboswitch aptamer forms a transcriptional terminator in the downstream expression platform, and this results in decreased levels of mRNAs encoding methionine biosynthesis enzymes and enzymes in other sulfur metabolism pathways that require SAM.
Conversely, some riboswitches that control transcription employ the opposite form of control—binding of the metabolite removes the stem–loop structure that terminates transcription. When this occurs, increased levels of the mature mRNA are eventually formed and, following translation, yield increased levels of the protein(s) it encodes.
Riboswitches and Evolution
How widespread are riboswitches and how did they evolve? Thus far riboswitches have been found only in some Bacteria, Archaea, and a few plants and fungi. Some scientists believe that riboswitches are remnants of the RNA world, a period eons ago before cells, DNA, and protein were present when it is hypothesized that catalytic RNAs were the only self-replicating “life forms” (Section 13.1 and Figure 13.4). In such an environment, riboswitches may have been an early mechanism of metabolic control—a simple means by which RNA life forms could have controlled the synthesis of other RNAs. As proteins evolved and DNA became the genetic material of all cells, riboswitches might have been the first control mechanisms for RNA (and even protein) synthesis as well. If this is true, the riboswitches that we see today may be the last vestiges of this simple form of control because, as we have seen in this chapter, metabolic regulatory mechanisms in cells are almost exclusively carried out by regulatory proteins.
We end this part of our study of microbial control with the description in the next section of one additional mechanism, a fascinating process by which cells essentially “decide” on whether to complete translation of a protein even before transcription of the mRNA that encodes it has been completed.
Check Your Understanding
What happens when a riboswitch binds the small metabolite that regulates it?
What are the major differences between a repressor protein and a riboswitch in the control of gene expression?
7.14 Attenuation
Attenuation is a form of transcriptional control in Bacteria and Archaea that functions by prematurely terminating mRNA synthesis. That is, in attenuation, control is exerted after the initiation of transcription but before its completion. Consequently, the number of completed transcripts from an operon is reduced, even though the number of initiated transcripts is not.
The basic principle of attenuation is that the first part of the mRNA to be made, called the leader, can fold into two alternative secondary structures. In this respect, the mechanism of attenuation resembles that of riboswitches that terminate transcription (Figure 7.31b). In attenuation, one mRNA secondary structure allows continued synthesis of the mRNA, whereas the other secondary structure causes premature termination. Folding of the mRNA depends either on events at the ribosome or on the activity of regulatory proteins, depending on the organism. The best examples of attenuation are the regulation of genes controlling the biosynthesis of certain amino acids in gram-negative Bacteria. The first to be described was in the tryptophan operon in Escherichia coli, and we focus on it here. Unlike the situation in prokaryotic cells, the processes of transcription and translation are spatially separated in eukaryotes (Section 6.6), and thus attenuation control is absent from Eukarya.
Attenuation in the Tryptophan Operon
The tryptophan operon contains structural genes for five proteins of the tryptophan biosynthetic pathway plus the usual promoter and regulatory sequences at the beginning of the operon (Figure 7.32). Like many operons, the tryptophan operon has more than one type of regulation. Transcription of the entire tryptophan operon is controlled by a repressor protein and its corepressor (tryptophan; Section 7.3). However, in addition to the promoter and operator regions needed for negative control, there is a sequence in the operon called the leader sequence that encodes a short polypeptide—the leader peptide—that functions as a translational barometer. The leader sequence contains tandem tryptophan codons near its terminus and is the “attenuator” of the attenuation system (Figure 7.32).
Figure 7.32 Attenuation and leader peptides in ***Escherichia coli***.

Structure of the tryptophan (trp) operon and of the tryptophan leader peptide and other leader peptides in E. coli. (a) Arrangement of the trp operon. Note that the leader (L) encodes a short peptide containing two tryptophan residues near its terminus (there is a stop codon following the Ser codon). The promoter is labeled P, and the operator is labeled O. The genes labeled trpE through trpA encode the enzymes needed for tryptophan synthesis. (b) Amino acid sequences of leader peptides of some other amino acid biosynthesis operons. Because isoleucine is made from threonine, it is an important constituent of the threonine leader peptide.
The basis of control of the tryptophan attenuator is as follows. If tryptophan is plentiful in the cell, there will be plenty of charged tryptophan tRNAs and the leader peptide will be synthesized. Synthesis of the leader peptide results in termination of transcription of the remainder of the trp operon, which includes the structural genes for the biosynthetic enzymes. By contrast, if tryptophan is scarce, the tryptophan-rich leader peptide will not be synthesized. If synthesis of the leader peptide is arrested by a shortage of tryptophan, the rest of the operon is transcribed.
Mechanism of Attenuation
How does translation of the leader peptide regulate transcription of the tryptophan genes downstream? Consider that in prokaryotic cells transcription and translation are simultaneous processes; as mRNA is released from the DNA, the ribosome binds to it and translation begins (Section 6.1 and Figure 6.6). That is, while transcription of downstream DNA sequences is still proceeding, translation of transcribed sequences is already under way (Figure 7.33).
Figure 7.33 Mechanism of attenuation.

Control of transcription of the Escherichia coli tryptophan (trp) operon structural genes by attenuation. The leader peptide is encoded by regions 1 and 2 of the mRNA. Two regions of the growing mRNA chain are able to form double-stranded loops, labeled as 3:4 and 2:3. (a) When there is excess tryptophan, the ribosome translates the complete leader peptide, and so region 2 cannot pair with region 3. Regions 3 and 4 then pair to form a loop that terminates transcription. (b) If translation is stalled because of tryptophan starvation, a loop forms by pairing of region 2 with region 3, loop 3:4 does not form, and transcription proceeds past the leader sequence.
Transcription is attenuated because a portion of the newly formed mRNA folds into a unique stem–loop that inhibits RNA polymerase activity. The stem–loop structure forms in the mRNA because two stretches of nucleotides near each other are complementary and can thus base-pair. If tryptophan is plentiful, the ribosome translates the leader sequence until it comes to the leader stop codon. The remainder of the leader sequence then forms a stem–loop, a transcription pause site, which is followed by a uracil-rich sequence that actually triggers termination (Figure 7.33a).
If the concentration of tryptophan in the cell is limiting, transcription of genes encoding tryptophan biosynthetic enzymes is needed. Thus, during transcription of the leader, the ribosome pauses at a tryptophan codon because of a shortage of charged tryptophan tRNAs. The presence of the stalled ribosome at this position allows a stem–loop to form (sites 2 and 3 in Figure 7.33b) that differs from the terminator stem–loop. This alternative stem–loop is not a transcription termination signal but instead prevents the terminator stem–loop (sites 3 and 4 in Figure 7.33a) from forming. This allows RNA polymerase to move past the termination site and begin transcription of tryptophan structural genes. Thus, in attenuation, we see a situation in which the rate of transcription is ultimately influenced by the rate of translation, a situation that contrasts with typical cellular events where the rate of translation is controlled by the rate of transcription.
We finish up this chapter on regulation by examining in Part V two major mechanisms that cells have evolved to control the activities of proteins they have already made.
Check Your Understanding
Why does attenuation control not occur in eukaryotes?
Explain how the formation of one stem–loop in the RNA can block the formation of another.
V Regulation of Enzymes and Other Proteins
Once a protein is made, its activity may still be subject to control. This ensures that cellular metabolites are synthesized in a balanced fashion and that energy is not expended making metabolites already present in excess of biosynthetic needs.
We have just explored some of the key mechanisms for regulating the amount (or even the complete presence or absence) of an enzyme or other protein within a cell. Here we focus on the mechanisms that cells employ to control the activity of enzymes already present in the cell through processes such as feedback inhibition and post-translational regulation.
7.15 Feedback Inhibition
A major means of controlling enzymatic activity is by feedback inhibition. This mechanism temporarily shuts off the reactions in an entire biosynthetic pathway. The reactions are shut off because an excess of the end product of the pathway inhibits activity of an early (typically the first) enzyme of the pathway. Inhibiting an early step effectively shuts down the entire pathway because no intermediates are generated for subsequent enzymes in the pathway (**Figure 7.34*a***). Feedback inhibition is reversible, however, because once levels of the end product become limiting, the pathway again becomes functional.
Figure 7.34 Inhibition of enzyme activity.

(a) In feedback inhibition, the activity of the first enzyme of the pathway is inhibited by the end product, thus shutting off the production of the three intermediates and the end product. (b) The mechanism of allosteric inhibition by the end product of a pathway. When the end product binds at the allosteric site, the conformation of the enzyme is so altered that the substrate can no longer bind to the active site. However, inhibition is reversible, and end product limitation will once again activate the enzyme. (c) Inhibition by isoenzymes. In Escherichia coli, the pathway leading to the synthesis of the aromatic amino acids contains three isoenzymes, each of which form a key intermediate, DAHP. Each isoenzyme is feedback-inhibited by a different aromatic amino acid. An excess of all three amino acids is required to completely shut off synthesis of DAHP and thus shut down the pathway.
How can the end product of a pathway inhibit the activity of an enzyme whose substrate is quite unrelated to it? This occurs because the inhibited enzyme has two binding sites, the active site (where substrate binds, Section 3.5), and the allosteric site, where the end product of the pathway binds. When the end product is in excess, it binds at the allosteric site, changing the conformation of the enzyme such that the substrate can no longer bind at the active site (Figure 7.34b). When the concentration of the end product in the cell begins to fall, however, the end product no longer binds to the allosteric site, so the enzyme returns to its catalytic form and once again becomes active.
Mastering Microbiology
Art Activity: Figure 7.34a, b Inhibition of enzyme activity
Some biosynthetic pathways controlled by feedback inhibition employ isoenzymes (“iso” means “same”). Isoenzymes are different proteins that catalyze the same reaction but are subject to different regulators, such as those required for the synthesis of the aromatic amino acids tyrosine, tryptophan, and phenylalanine in Escherichia coli, a branched biosynthetic pathway (Figure 7.34c). Three isoenzymes catalyze formation of DAHP, an early intermediate of the pathway, and each is regulated independently by a different end product amino acid. In this way, synthesis of DAHP is inhibited incrementally, an excess of all three amino acids being necessary to completely shut down the pathway. This form of feedback inhibition prevents the excess of one or two of the amino acids from shutting down the entire pathway and thereby starving the cell for the third amino acid (Figure 7.34c).
Check Your Understanding
What is the difference between an allosteric site and an active site?
7.16 Post-Translational Regulation
We have already discussed phosphorylation and methylation, two very common mechanisms for regulating a protein post-translationally, when we considered two-component regulatory systems and chemotaxis, respectively (Sections 7.5 and 7.6). Biosynthetic enzymes can also be regulated by the attachment of other small molecules, such as the nucleotides adenosine monophosphate (AMP), adenosine diphosphate (ADP), and uridine monophosphate (UMP). Throughout this chapter we have also encountered avenues in which the cell regulates the activity of proteins by protein–protein interactions. In contrast to these, some enzymes are regulated by covalent modification, typically the attachment or removal of a small molecule to or from the enzyme that subsequently affects its catalytic activity, and we consider this regulatory mechanism here.
Regulation of PII Signal Transduction Proteins
PII proteins are a widespread family of signal-transducing proteins (Section 7.5) found in Bacteria, Archaea, and the plastids of plants. PII proteins play an important role in nitrogen metabolism by regulating a diverse range of transcription factors, enzymes, and membrane transport proteins (we have already seen one instance of PII-based regulation with the Ntr system, Figure 7.14b). The activity of PII proteins on their regulatory cascade depends on whether they have been covalently modified or not. These modifications range from uridylylation (addition of a UMP group, Figure 7.14b), adenylylation (addition of AMP), or even phosphorylation in some organisms. Because these covalent modifications affect protein activity and occur after the PII proteins have been fully synthesized, the regulation is post-translational. Here we consider PII and its uridylylation by GlnD, a bifunctional enzyme that possesses both uridylyltransferase activity and uridyl-removal activity in response to levels of the amino acid glutamine (Figure 7.35).
Figure 7.35 Post-translational regulation of PII and glutamine synthetase.

Glutamine synthetase is a key enzyme of ammonia incorporation, especially when ammonia is present at low concentrations, in all Bacteria and Archaea (Section 3.14). (a) Uridylylation of PII and activation of glutamine synthetase (GS), a multi-subunit enzyme. In response to low cellular pools of glutamine, the uridylyltransferase activity of GlnD is stimulated. This converts PII to PII-UMP, which triggers glutamine synthetase adenyltransferase to activate GS by removing AMP(s). (b) Removal of UMP from PII and inactivation of GS. In response to high cellular pools of glutamine, the UMP removal activity of GlnD is activated. This converts PII-UMP to PII, which triggers glutamine synthetase adenyltransferase to inactivate GS through adenylylation. (c) Overall control of GS activity. Adenylylated GS subunits are catalytically inactive, so the overall GS activity increases progressively as glutamine levels decrease and more subunits are deadenylylated. Conversely, GS activity decreases progressively as glutamine levels increase and more subunits are adenylylated.
One of the main ways the cell determines if ammonia (NH3) assimilation is necessary is through glutamine sensing by GlnD. If the cellular glutamine pool is low, it signals that NH3 assimilation is necessary. GlnD transduces this signal through the binding of glutamine itself. When GlnD is not bound to glutamine, the enzyme possesses uridylyltransferase activity and adds a UMP group to PII, yielding PII-UMP by post-translational modification.
One protein target of PII-UMP within the nitrogen metabolism pathway is glutamine synthetase adenyltransferase (Figure 7.35a). Once uridylylated, PII-UMP stimulates glutamine synthetase adenyltransferase to remove adenyl groups from glutamine synthetase (GS). GS is a key energy-requiring enzyme in ammonia (NH3) assimilation (Section 3.14) and must be tightly regulated to conserve energy if cellular levels of nitrogen are high, as sensed by the glutamine pool. As GS becomes less adenylylated, its activity increases and NH3 assimilation increases (Figure 7.35c). Once glutamine levels are sufficient, the cell no longer needs to assimilate NH3. This results in GlnD binding to available glutamine, which stimulates the protein to remove uridyl groups from PII-UMP (Figure 7.35b). PII in its unmodified form stimulates glutamine synthetase adenyltransferase to adenylate GS, forming GS-AMP. Fully adenylylated GS is less active and thus NH3 assimilation by way of GS diminishes.
Why does all this elaborate regulation surround the enzyme GS? It is basically a matter of conserving energy. Because it has such a high affinity for its substrate, GS can assimilate ammonia present at very low levels, but this capability comes at a cost—the expenditure of ATP. However, when NH3 is present at high levels in the cell, it can be assimilated into amino acids by enzymes that do not consume ATP; under these conditions, GS in the cell remains inactive. Then, when NH3 levels are low, GS activity increases. By having GS active only when NH3 is limiting, the cell conserves ATP that could be used to drive other energy-consuming processes in the cell.
Inactivation of Sigma Factors
In Section 7.11 we described how the σ factor RpoH is inactivated by DnaK under normal temperature conditions in the heat shock response (Figure 7.27). Proteins known as anti-sigma factors can also bind to sigma factors, inactivating them in a form of post-translational regulation.
RpoE (σ24 or σE, Table 6.3) is a sigma factor widespread in Bacteria that responds to extracytoplasmic stress by recognizing promoters for genes that encode products necessary for proper folding, expression, and turnover of proteins that reside in the outer membrane (Section 2.4). One of these promoters is for dnaK. Under nonstressful conditions, the membrane-bound anti-sigma factor RseA sequesters RpoE from binding to the promoters it targets (**Figure 7.36*a***). However, if a cell encounters a condition stressful to the membrane such as heat or osmotic stress, outer membrane proteins are denatured, and this activates a protease specific for RseA (Figure 7.36b). Once RseA has been degraded, RpoE is free to activate transcription of genes necessary for cell envelope repair such as refolding outer membrane proteins and the synthesis of phospholipids and lipopolysaccharides. Release of RpoE also includes increasing the level of DnaK produced so that this chaperone (Section 6.11) can assist with folding and turnover of outer membrane proteins as well.
Figure 7.36 Anti-sigma–sigma factor interactions.

(a) Sequestration of the RpoE sigma factor by the anti-sigma factor RseA. When the outer membrane is not stressed, cytoplasmic membrane–bound RseA binds to RpoE. This binding prevents the sigma factor from binding to promoters of genes for envelope repair and dnaK. (b) Inactivation of RseA and release of the RpoE sigma factor. When the outer membrane is exposed to stress, outer membrane proteins (OMPs) are denatured. These unfolded OMPs trigger proteolytic degradation of RseA. Without RseA, the RpoE sigma factor can bind to its promoter regions and transcribe genes for envelope stress and dnaK to help repair the membrane and refold proteins.
Once the membrane has been repaired, RseA is no longer targeted for degradation and reassociates with the cytoplasmic membrane. In this form, RseA can resume its anti-sigma factor activity by binding and sequestering RpoE from promoter regions. This leads to a decreased expression of both genes encoding products needed to respond to membrane stress and dnaK. Another well-studied example of regulation by anti-sigma–sigma interactions occurs during the formation of endospores in Bacillus when the anti-sigma factor SpoIIAB binds to σF, thereby preventing its association with RNA polymerase; we pick up on this control process in Section 8.6 and Figure 8.18.
Regardless of the mechanism, in the final analysis it should be clear that regulating the synthesis and activities of a cell’s RNAs and proteins is (1) very important to its biology, (2) possible in many different ways, and (3) a major genetic investment. But the costs to the cell are worth it and are grounded in billions of years of evolutionary refinement. At every turn in a highly competitive world, a microbe’s growth and very survival may well depend on its ability to conserve its resources and maximize its progeny.
Check Your Understanding
What types of covalent modifications commonly alter the activity of proteins?
How does GlnD signal to the cell that NH3 assimilation is necessary?
Chapter Review
Go to Mastering Microbiology for videos, animations, practice tests, and more.
I DNA-Binding Proteins and Transcriptional Regulation
7.1 Proteins can bind to DNA when specific protein domains bind to specific regions of the DNA molecule. In most cases the interactions are sequence-specific.
Q Explain why a protein that binds to a specific sequence of double-stranded DNA is unlikely to bind to the same sequence if the DNA is single-stranded.
7.2 Transcription factors are DNA-binding proteins that control gene expression. Activator and repressor proteins are common transcription factors whose DNA binding is controlled by the presence or absence of effector molecules.
7.3 The amount of a specific enzyme in the cell can be controlled by regulatory proteins that bind to DNA and increase (induce) or decrease (repress) the amount of mRNA that encodes the enzyme.
Q Why is enzyme induction considered the opposite of enzyme repression? What is the difference between an operon and a regulon?
7.4 Archaea resemble Bacteria in using DNA-binding activator and repressor proteins to regulate gene expression at the level of transcription.
Q What are the two mechanisms used by archaeal repressor proteins to repress transcription?
II Sensing and Signal Transduction
7.5 Signal transduction systems transmit environmental signals to the cell. In Bacteria and Archaea, signal transduction is typically carried out by a two-component regulatory system that includes a membrane-integrated sensor kinase and a cytoplasmic response regulator. The activity of the response regulator depends on its state of phosphorylation.
Q What are the two components that give their name to the signal transduction system in prokaryotic cells? What is the function of each of the components?
7.6 Chemotactic behavior allows cells to respond to attractants and repellents in their environment. The regulation of chemotaxis affects the activity of proteins rather than their synthesis. Adaptation by methylation allows the system to reset itself to the continued presence of a signal.
Q Adaptation allows the mechanism controlling flagellar rotation to be reset. How is this achieved?
7.7 Quorum sensing allows cells to monitor their environment for cells of their own kind. Quorum sensing depends on the sharing of specific small molecules known as autoinducers. Once a sufficient concentration of the autoinducer is present, specific gene expression is triggered.
Q How can quorum sensing be considered a regulatory mechanism for conserving cell resources?
III Global Control
7.8 Global control systems regulate the expression of many genes simultaneously. Catabolite repression is a global control system that helps cells make the most efficient use of available carbon sources. The lac operon is under the control of catabolite repression as well as its own specific negative regulatory system.
Q Describe the mechanism by which cAMP receptor protein (CRP), the regulatory protein for catabolite repression, functions. Use the lactose operon as an example.
7.9 The stringent and general stress responses are deployed to survive nutrient limitation and stresses by decreasing expression of genes for macromolecule biosynthesis and activating stress survival pathways.
**Q Explain the sequence of molecular events that leads to the synthesis of (p)ppGpp in Escherichia coli during the stringent response.**
7.10 The phosphate (Pho) regulon employs a two-component regulatory system to sense cellular levels of phosphate. The cytoplasmic transcriptional regulator of this regulon binds to promoter regions called Pho boxes and functions as either an activator or a repressor depending on phosphate levels in the cell.
Q Explain how the binding of Pho transcriptional regulator activates transcription of some genes within the Pho regulon, but not others.
7.11 Cells respond to excessive temperature and other stressors that lead to protein denaturation by employing the heat shock response. This response results in expressing sets of genes whose products help the cell overcome stress, including an alternative sigma factor.
**Q Describe the proteins produced when cells of Escherichia coli experience a heat shock. Of what value are they to the cell?**
IV RNA-Based Regulation
7.12 Cells can control genes in several ways by employing regulatory RNA molecules. One way is to take advantage of base pairing and use small RNAs to promote or prevent translation of mRNAs.
Q What are the mechanisms by which regulation by sRNA occurs?
7.13 Riboswitches are sequences at the 5′ ends of certain mRNAs that recognize small molecules and respond by changing their three-dimensional structure to affect translation or transcriptional termination of the mRNA. Riboswitches are mostly employed to control biosynthetic pathways for vitamins, amino acids, purines, and a few other metabolites.
Q What are the mechanisms by which riboswitches regulate translation and transcription?
7.14 Attenuation is a mechanism whereby transcription is controlled after initiation of mRNA synthesis. Attenuation mechanisms depend upon alternative stem–loop structures in the mRNA that result in either read-through or stalling of the ribosome.
Q Describe how transcriptional attenuation works. What is actually being “attenuated”?
V Regulation of Enzymes and Other Proteins
7.15 In feedback inhibition, an excess of the final product of a biosynthetic pathway inhibits an allosteric enzyme at the beginning of the pathway. Enzyme activity can also be modulated by isoenzymes.
Q Describe how feedback inhibition is reversible, and why are isoenzymes necessary for some forms of feedback inhibition?
7.16 Protein activity can be regulated after translation. Reversible covalent modification or interactions with other proteins can modulate protein activity.
Q Which nucleotides are commonly used to covalently modify protein activity?
Application Questions
What would happen to regulation from a promoter under repression if the region where the regulatory protein binds was deleted? What if the promoter was under positive control? Promoters from Escherichia coli under positive control are not close matches to the promoter consensus sequence for E. coli. Why?
Most of the regulatory systems described in this chapter employ regulatory proteins. However, regulatory RNA is also important. Describe how one could achieve negative control of the lac operon using either of two different types of regulatory RNA.
Many amino acid biosynthetic operons under attenuation control are also under negative control. Considering that the environment of a bacterium can be highly dynamic, what advantage could be conferred by having attenuation as a second layer of control?
How would you design a regulatory system to make Escherichia coli use succinic acid in preference to glucose? How could you modify it so that E. coli prefers to use succinic acid in the light but glucose in the dark?
Chapter Glossary
a regulatory protein that binds to specific sites on DNA and stimulates transcription Allosteric protein
a protein containing an active site for binding substrate and an allosteric site for binding an effector molecule such as the end product of a biochemical pathway Attenuation
a mechanism for controlling gene expression that terminates transcription after initiation but before a full-length messenger RNA is produced Autoinducer
a small signal molecule that takes part in quorum sensing Catabolite repression
the suppression of alternative catabolic pathways by a preferred source of carbon and energy Cyclic AMP
a regulatory nucleotide that participates in catabolite repression Domain
region of a protein with specific structure and function Enzyme induction
production of an enzyme in response to a signal (often the presence of the substrate for the enzyme) Enzyme repression
prevention of the synthesis of an enzyme in response to a signal Feedback inhibition
a process in which an excess of the end product of a multistep pathway inhibits activity of the first enzyme in the pathway Gene expression
transcription of a gene followed by translation of the resulting mRNA into protein Heat shock proteins
proteins induced by high temperature (or certain other stresses) that protect against high temperature, especially by refolding partially denatured proteins or by degrading them Heat shock response
response to high temperature that includes the synthesis of heat shock proteins together with other changes in gene expression Noncoding RNA (ncRNA)
RNA that is not translated into protein; examples include ribosomal RNA, transfer RNA, and small regulatory RNAs Operon
two or more genes transcribed into a single RNA and under the control of a single regulatory site Quorum sensing
a regulatory system that monitors the population level and controls gene expression based on cell density Regulatory nucleotide
a nucleotide that functions as a signal rather than being incorporated into RNA or DNA Regulon
a series of operons controlled as a unit Repressor protein
a regulatory protein that binds to specific sites on DNA and blocks transcription Response regulator protein
one of the members of a two-component regulatory system; a protein that is phosphorylated by a sensor kinase and then acts as a regulator, often by binding to DNA Riboswitch
an RNA domain, usually in a messenger RNA molecule, that can bind a specific small molecule, altering its own secondary structure; this, in turn, controls translation of the mRNA Sensor kinase protein
one of the members of a two-component regulatory system; a protein that phosphorylates itself in response to an external signal and then transfers the phosphoryl group to a response regulator protein Signal transduction
see two-component regulatory system Stringent response
a regulatory mechanism that detects nutrient or environmental stresses and antibiotics Transcription factor
a regulatory protein that binds to specific sites on DNA and controls transcription; generally activator and repressor proteins Two-component regulatory system
a regulatory system consisting of two proteins: a sensor kinase and a response regulator