9 Genetics of Bacteria and Archaea
**III Gene Transfer in Archaea and Other Genetic Events**
Live Cell Imaging Captures Bacterial Promiscuity
While often considered asexual, both Bacteria and Archaea evolve quickly through DNA exchange by horizontal transfer. Increased fitness resulting from genetic promiscuity has traditionally been monitored in the laboratory through changes in physiology. However, advances in microscopy and fluorescent labeling now make it possible to witness DNA exchange using live cell imaging.
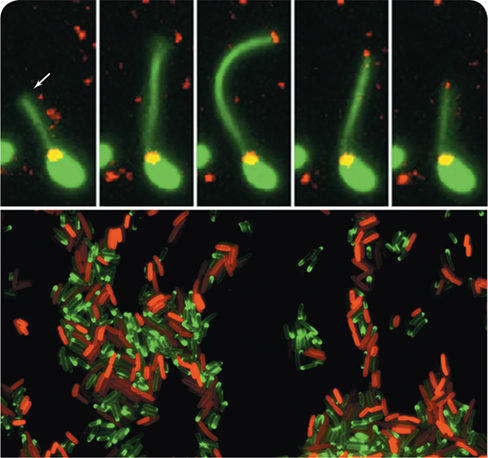
One fascinating example is the DNA uptake system (natural competence) in the bacterium Vibrio cholerae. Using an appendage called a pilus, V. cholerae is able to reach outside of its cell, snatch pieces of free DNA, and transport them into the cell. Thus, its pili effectively function as “harpoons” fishing for free DNA. The top panel of the images shown here is a time series of the “harpoon” (white arrow) being cast from the cell into the environment, attaching to free DNA (red), and then retracting back to the cell.1
Transfer of antibiotic resistance can also be monitored using live cell imaging. The photo at the bottom shows a mixed population of Escherichia coli cells surrounded by the antibiotic tetracycline (labeled green). Cells possessing a plasmid encoding resistance to tetracycline appear red due to the labeled TetA resistance protein, whereas cells sensitive to tetracycline appear green due to their uptake of the labeled antibiotic.2 Why are the sensitive cells still alive? Remarkably, an efflux pump (Chapter 8) helps the sensitive cells buy just enough time to obtain the plasmid from their resistant neighbors through a genetic event called conjugation. How long does this whole process take? Transfer of the plasmid only takes minutes, and sensitive cells can express TetA in under 2 hours.
These examples highlight the astounding power of horizontal gene transfer in prokaryotic cells as well as the variety of tools at their disposal to improve their fitness. Bacterial genetics is indeed an exciting and dynamic discipline and has given birth to many of the genetic concepts we know today.
Sources: 1Ellison, C.K., et al. 2018. Retraction of DNA-bound type IV competence pili initiates DNA uptake during natural transformation of Vibrio cholerae. Nat. Microbiol. 3: 773.
2Nolivos, S., et al. 2019. Role of AcrAB-TolC multidrug efflux pump in drug-resistance acquisition by plasmid transfer. Science 364: 778.
The diversity and amazing ability of bacteria to survive ever-changing environmental conditions and a host of stressors is controlled by their genetic makeup. While their asexual reproduction would suggest that cells within Bacteria and Archaea do not exchange genes, the microbiologist Joshua Lederberg made a groundbreaking discovery in 1946—like plants and animals, bacteria can also exchange genes! Lederberg’s groundbreaking work showcasing genetic recombination in Bacteria not only earned him a Nobel Prize but also helped launch the field of molecular biology and the use of Bacteria to study how genes work in higher organisms, such as plants and animals.
Understanding the varied mechanisms by which Bacteria and Archaea exchange genes has helped tackle the conundrum of how these microbes can exhibit so much diversity and persist in habitats where other forms of life cannot. Gene exchange between microbes, along with genetic innovations that arise from random changes in a cell’s genetic blueprint, occur frequently and confer selectable advantages that ultimately drive cell survival and genetic diversity.
In this chapter we discuss the major themes of bacterial and archaeal genetics and the processes of gene transfer from one cell to another (Figure 9.1). We first describe how changes arise in the genome, which can be due to random errors in DNA replication or from DNA damage, and then we consider how horizontal gene transfer (Figure 9.1; see also the photomicrograph of actual gene transfer on page 261) moves genes from one cell to another by mechanisms uncoupled from reproduction. Microbiologists have leveraged aspects of these mutational and genetic exchange processes to identify genes encoding exciting microbial processes such as chemotaxis, quorum sensing, and persistence that we discussed in Chapters 7 and 8. While changes to the genome underlie microbial diversity and habitat adaptation, microorganisms also possess mechanisms to maintain genomic stability, and we end this chapter by considering these. Taken together, both genomic change and genomic stability are important to the evolution of an organism and its competitive success in nature.
Figure 9.1 Overview of bacterial and archaeal genetics.

DNA mutations and mechanisms of DNA transfer contribute to the genetic diversity of Bacteria and Archaea. Transduction, transformation, and conjugation are the three known ways by which prokaryotic cells can exchange genes.
I Mutation
Changes in the human world can be good, bad, or indifferent, and the same is true of the microbial world. Changes in the genetic blueprint of a microbe may fuel its evolutionary progress, accelerate its death, or have no detectable effect on its biology.
All organisms contain a specific sequence of nucleotides in their genome, their genetic blueprint. A mutation is a heritable change in the base sequence of that genome, that is, a change that is passed from the mother cell to progeny cells. Mutations can lead to changes in the properties of an organism; some mutations are beneficial, some are detrimental, but most are neutral and have no effect. Although the rate of spontaneous mutation is low (Section 9.3), the impressive characteristic of exponential growth in Bacteria and Archaea ensures that mutations accumulate in a population surprisingly fast. Moreover, whereas a single mutation typically brings about only a small change in a cell, horizontal gene transfer often generates much larger changes. Taken together, mutation and genetic exchange fuel the evolutionary process.
We begin by considering the molecular mechanism of mutation and the properties of mutant microorganisms.
9.1 Mutations and Mutants
In order to appreciate the impact that genome changes can have on a cell (or a virus), we start with a brief genetics primer. The genomes of cells consist of double-stranded DNA. In viruses, by contrast, the genome may consist of double- or single-stranded DNA or RNA, depending on the virus (Chapters 5 and 11). By convention, a strain of an organism or a virus isolated from nature is called the wild-type strain and therefore contains the wild-type genome. A cell or virus derived from the wild type whose genome carries a change in nucleotide sequence is called a mutant. A mutant by definition differs from the wild-type strain in its genotype, the nucleotide sequence of its genome (**Figure 9.2*a***). In addition, the observable properties of a mutant—its phenotype—may also be altered relative to its parent (Figure 9.2b). This altered phenotype is called a mutant phenotype.
Figure 9.2 Wild-type versus mutant phenotype.

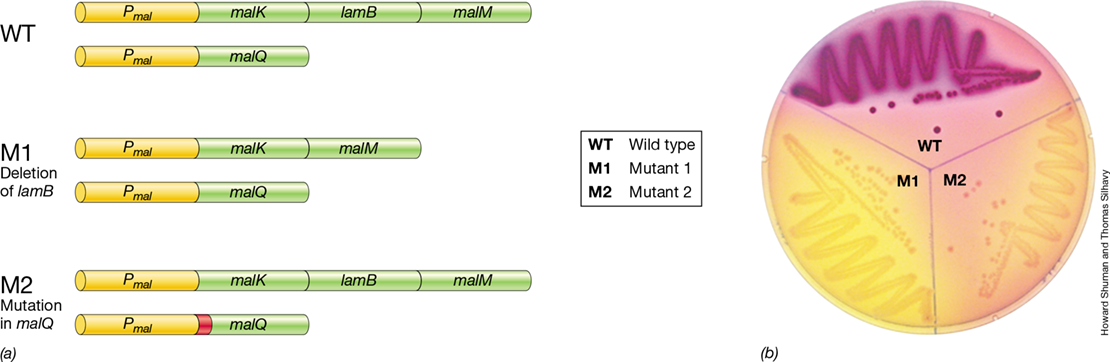
(a) Genotype maps of Escherichia coli wild-type and maltose utilization mutants; P is the promoter region and mal and lam regions are structural genes. (b) Growth of wild-type E. coli and mutants on a plate of MacConkey agar, a differential medium. The medium contains maltose as the carbon source and a pH indicator that turns red if maltose is fermented. While the wild-type strain ferments maltose, mutant strains M1 and M2 are unable to ferment maltose due to a deletion of the lamB gene and a point mutation in the malQ gene, respectively.
The term “wild type” may be used to refer to an entire organism or just to the status of a particular gene that is under investigation. Mutant derivatives can be obtained either directly from a wild-type strain or from another strain—referred to as a parental strain—previously derived from the wild type; for example, another mutant. Figure 9.2b shows a plate of MacConkey agar (a culture medium that contains a pH indicator that turns red if sugar is fermented) containing the sugar maltose. This medium shows the phenotypic difference between wild-type Escherichia coli and mutant derivatives in the maltose utilization pathway.
Depending on the mutation, a mutant strain may or may not differ in phenotype from its parent. By convention in bacterial genetics, the genotype of an organism is designated by three lowercase letters followed by a capital letter (all in italics) indicating a particular gene. For example, the hisC gene of E. coli encodes a protein called HisC that functions in biosynthesis of the amino acid histidine (Figure 6.27). Mutations in the hisC gene would be designated as hisC1, hisC2, and so on, the numbers referring to the order of isolation of the mutant strains. Each hisC mutation would be different, and each hisC mutation might affect the HisC protein in different ways.
The phenotype of an organism is designated by a capital letter followed by two lowercase letters, with either a plus or a minus superscript to indicate the presence or absence of that property. For example, a His+ strain of E. coli is one that is capable of making its own histidine, whereas a His− strain is not. The His− strain would therefore require a histidine supplement for growth. A mutation in the hisC gene leads to a His− phenotype if it eliminates the function of the HisC protein. Likewise, the lamB and malQ mutations in Figure 9.2 lead to a Mal− phenotype, as the mutants can no longer use maltose as an energy source.
Isolation of Mutants: Screening versus Selection
Virtually any characteristic of an organism can be changed by mutation. Some mutations are selectable, conferring some type of advantage on organisms possessing them, whereas others are nonselectable, even though they may lead to a very clear change in the phenotype of an organism. A selectable mutation confers a clear advantage on the mutant strain under certain environmental conditions, so the progeny of the mutant cell are able to grow and replace the parent. A good example of a selectable mutation is drug resistance: An antibiotic-resistant mutant can grow in the presence of an antibiotic that inhibits or kills the parent (**Figure 9.3*a***) and is thus selected under these conditions. It is relatively easy to detect and isolate selectable mutants by choosing the appropriate environmental conditions. Selection is therefore an extremely powerful genetic tool, allowing the isolation of a single mutant from a population containing millions or even billions of parental cells.
Figure 9.3 Selectable and nonselectable mutations.
(a) Development of antibiotic-resistant mutants, a type of easily selectable mutation, within the inhibition zone of an antibiotic assay disk. (b) Nonselectable mutations. UV-radiation-induced nonpigmented mutants of Serratia marcescens. The wild type has a dark red pigment. The white or colorless mutants make no pigment. (c) Colonies of mutants of a species of Halobacterium, a member of the Archaea. The lower pink wild-type colony contains intracellular gas vesicle nanoparticles that regulate buoyancy and also scatter light, resulting in a milky appearance. The upper red-orange colony is a mutant lacking gas vesicles and is translucent as a result.
Some examples of important nonselectable mutations are secondary metabolite production loss in an antibiotic-producing organism, loss of attachment ability in a biofilm-forming organism, and loss of color in a pigmented organism (Figure 9.3b, c). All three types of mutants usually have neither an advantage nor a disadvantage over their parent cells when grown in the laboratory, although antibiotic production (Section 16.12), biofilm formation (Sections 4.9 and 8.10, and Section 20.4), and pigmentation (for example, of phototrophic organisms) may have a selective advantage in nature. We can detect nonselectable mutations only by examining large numbers of colonies and looking for the “different” ones, an often laborious process called screening.
Mastering Microbiology
Art Activity: Figure 9.4 Screening for nutritional auxotrophs
As illustrated in Figure 9.2 (see also Figure 9.35), a thorough understanding of microbial physiology is the key to designing successful genetic screens for nonselectable mutations. In Figure 9.2b a genetic screen for Mal− mutants utilizes a culture medium containing a pH indicator that turns red if maltose is fermented (MacConkey agar). Because the Mal− mutants are unable to utilize maltose, they are unpigmented compared to the red wild-type strain. To identify mutants in biofilm formation, individual colonies can be grown in liquid medium in a microtiter plate and a stain can be used to track whether cells can form a biofilm and adhere to the microtiter plate (see Figure 9.35). If possible, selection is almost always the preferred strategy over screening for obtaining mutants in a genetic experiment because selective conditions typically place such severe restraints on the population that mutants are easily detectable. Examples of common classes of mutants and the means by which they are detected are listed in Table 9.1.
Isolation of Nutritional Auxotrophs
Although screening is more tedious than selection, useful methods have been developed for screening large numbers of colonies for certain types of mutations. For instance, nutritionally defective mutants can be detected by the technique of replica plating (Figure 9.4). A colony from a master plate can be transferred onto an agar plate lacking the nutrient by using a sterile loop, toothpick, or even a robotic arm. Parental colonies will grow normally, whereas those of the mutant will not. Thus, the inability of a colony to grow on a medium lacking the nutrient signals that it is a mutant. The colony on the master plate corresponding to the vacant spot on the replica plate can then be picked, purified, and characterized.
Figure 9.4 Screening for nutritional auxotrophs.

The replica-plating method can be used for the detection of nutritional mutants. Colonies from the master plate are transferred using a sterile toothpick to a gridded plate containing different media for selection. The colonies not appearing on the selective medium are labeled as auxotrophs. The selective medium lacked one nutrient (the amino acid leucine) present in the master plate. Therefore, the colonies on the complete medium plate that are not represented on the selective medium plate are leucine auxotrophs (Leu−).
A mutant strain with an additional nutritional requirement above that of the wild type or parental strain from which it was derived is called an auxotroph (Table 9.1), and the strain from which an auxotroph originates is called a prototroph. For instance, mutants of E. coli with His− and Mal− (Figure 9.2) phenotypes are histidine and maltose auxotrophs, respectively, while the parental His+ and Mal+ strains from which the auxotrophs were derived are the prototrophs of such strains. As described earlier, many different mutations can lead to a strain showing a His− or Mal− phenotype, and thus an initial step in characterizing the genetics of a metabolic pathway (such as histidine biosynthesis and maltose catabolism) would be the isolation of several His− or Mal− strains followed by their comparative genetic analyses (Figure 9.2). This comparative analysis process, called complementation, is discussed in Section 9.5.
9.2 Molecular Basis of Mutation
Mutations can be either spontaneous or induced events. Spontaneous mutations are those that occur without external intervention, and most result from occasional errors in the pairing of bases by DNA polymerase during DNA replication (Chapter 6). Induced mutations, by contrast, are those caused by environmental agents and include mutations made deliberately by humans. Induced mutations can result from exposure to natural radiation (cosmic rays and so on) that alters the structure of bases in the DNA, or from a variety of chemicals that chemically modify DNA (Section 9.4).
Mutations that change only one base pair are called point mutations and occur when a single base-pair substitution occurs in the DNA. Many point mutations do not actually cause any phenotypic change (although the organism’s genotype has changed). However, as for all mutations, any phenotypic change that results from a point mutation depends on exactly where in the genome the mutation occurs and the nature of the nucleotide change, as we will see.
Base-Pair Substitutions: Missense, Nonsense, and Silent Mutations
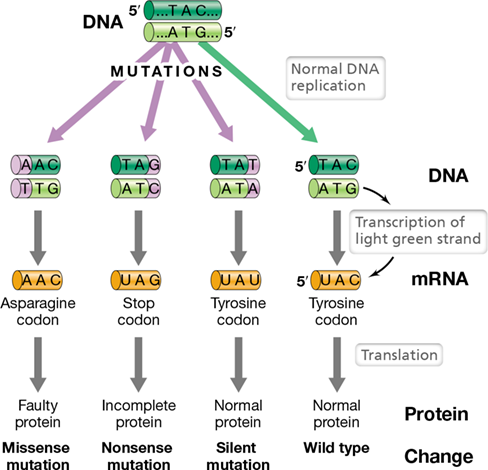
If a point mutation is within the region of a gene that encodes a polypeptide, any change in the phenotype of the cell is most likely the result of a change in the amino acid sequence of that polypeptide. The error in the DNA is transcribed into mRNA, and the erroneous mRNA in turn is translated to yield a polypeptide. In interpreting the results of a mutation, we must first recall that the genetic code is degenerate (Section 6.9 and Table 6.4). Consequently, not all mutations in the base sequence encoding a polypeptide will change the polypeptide sequence. This is illustrated in Figure 9.5, which shows several possible results when the DNA that encodes a single tyrosine codon in a polypeptide is mutated. First, a change in the RNA from UAC to UAU would have no apparent effect because UAU is also a tyrosine codon. Although they do not affect the sequence of the encoded polypeptide, such changes in the DNA are considered one type of silent mutation, that is, a mutation that does not affect the phenotype of the cell. Note that silent mutations in coding regions are almost always in the third base of the codon (arginine and leucine can also have silent mutations in the first position) because of genetic code degeneracy.
Figure 9.5 Possible effects of base-pair substitution in a gene encoding a protein.

Three different protein products are possible from changes in the DNA for a single codon. The possible phenotypes are wild type and strains harboring missense, nonsense, or silent mutations.
Changes in the first or second base of the codon more often lead to significant changes in the amino acid sequence of the polypeptide. For instance, a single base change from UAC to AAC (Figure 9.5) results in an amino acid change within the polypeptide from tyrosine to asparagine at a specific site. This is called a missense mutation because the informational “sense” (the precise sequence of amino acids) in the polypeptide has changed. If the change is at a critical location in the polypeptide chain, the protein could be inactive, have reduced activity, or have modified activity. However, not all missense mutations lead to nonfunctional proteins. The outcome depends on where the substitution lies in the polypeptide chain and on how it affects protein folding and activity. While mutations in the active site of an enzyme are more likely to destroy catalytic activity than mutations in other regions of the protein, some active site mutations can alter an enzyme’s substrate specificity. This type of change can result in a mutant enzyme with differing activity from the wild-type enzyme, and such events have contributed to the spectacular metabolic diversity we see in the microbial world (Chapters 3 and 14).
Another possible outcome of a base-pair substitution is the formation of a stop (nonsense) codon in the DNA. This results in premature termination of translation, leading to an incomplete polypeptide (Figure 9.5). Mutations of this type are called nonsense mutations because the change is from a sense (coding) codon to a nonsense (stop) codon (Table 6.4). Unless the nonsense mutation is very near the end of the gene, the product is incompletely made; such truncated proteins are either inactive or, at the very least, lack normal activity.
Other terms are occasionally used in microbial genetics to describe the precise type of base substitution in a point mutation. Transitions are mutations in which one purine base (A or G) is substituted for another purine, or one pyrimidine base (C or T) is substituted for another pyrimidine. Transversions are point mutations in which a purine base is substituted for a pyrimidine base, or vice versa.
Frameshifts and Other Insertions or Deletions
Because the genetic code is read from one end of the nucleic acid in consecutive blocks of three bases (codons), any deletion or insertion of a single base pair results in a shift in the reading frame. These frameshift mutations often have serious consequences. Single base insertions or deletions change the primary sequence of the encoded polypeptide, typically in a major way (Figure 9.6). Such microinsertions or microdeletions can result from DNA polymerase replication errors. Insertion or deletion of two base pairs also causes a frameshift. However, insertion or deletion of three base pairs does not cause a frameshift but does add or remove a codon; this results in the addition or deletion of a single amino acid in the polypeptide sequence. Although an amino acid addition or deletion may well be deleterious to protein function, it is usually not as serious a problem as a frameshift, which scrambles the entire polypeptide sequence downstream of the mutation.
Figure 9.6 Shifts in the reading frame of mRNA caused by insertions or deletions.

The reading frame in mRNA is established by properly positioning the message on the ribosome. The mRNA is read beginning at the 5′ end (toward the left in the figure) and proceeds by units of three bases (codons). The normal reading frame is referred to as the 0 frame, that missing a base the −1 frame, and that with an extra base the +1 frame.
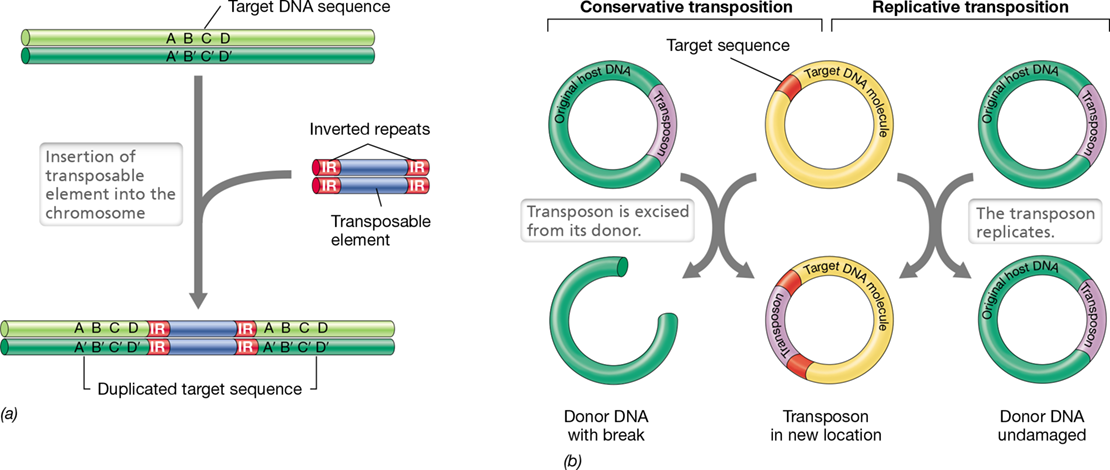
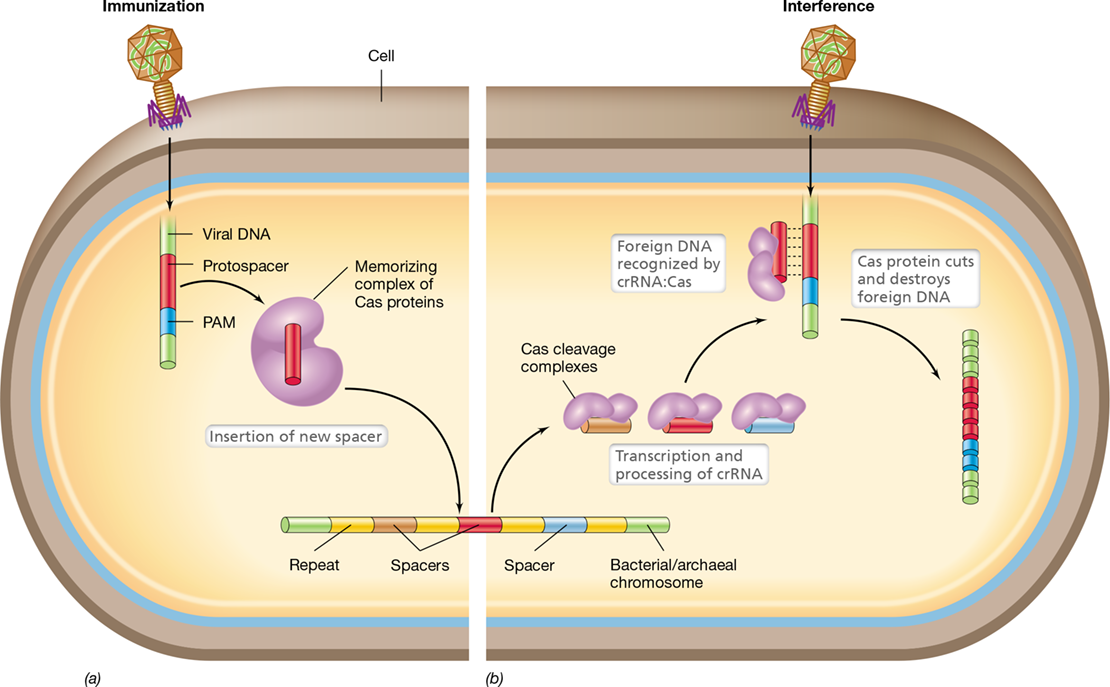
Insertions or deletions can also result in the gain or loss of hundreds or even thousands of base pairs. Such changes inevitably result in complete loss of gene function. Some deletions are so large that they may include several genes. If any of the deleted genes are essential, the mutation will be lethal. Such deletions cannot be restored through further mutations, but only through genetic recombination (Sections 9.5, 9.6, 9.7, 9.8, 9.9, 9.10 and 9.11). Large insertions and deletions may arise as a result of errors during genetic recombination. In addition, many large insertion mutations are due to the insertion of specific DNA sequences called transposable elements (Section 9.11).
We will discuss the effects of mutations on the evolution of bacterial genomes in Chapter 13.
Check Your Understanding
Do missense mutations occur in genes encoding tRNA? Why or why not?
Why do frameshift mutations generally have more serious consequences than missense mutations?
9.3 Reversions and Mutation Rates
The rates at which the different kinds of mutations occur vary widely. Some mutations occur so rarely that they are almost impossible to detect, whereas others occur so frequently that they present difficulties for an experimenter trying to maintain a genetically stable stock culture. Sometimes a second mutation can reverse the effect of an initial mutation. Furthermore, all organisms possess a variety of systems for DNA repair. Consequently, the observed mutation rate depends not only on the frequency of DNA changes but also on the efficiency of DNA repair.
Reversions (Back Mutations) and Suppressors
Point mutations are typically reversible, a process known as reversion. A revertant is therefore a strain in which the original phenotype that was changed by mutation is restored by a second mutation. Revertants can be of two types, same site or second site. In same-site revertants, the mutation that restores activity is at the same site as the original mutation. If the back mutation is not only at the same site but also restores the original sequence, it is called a true revertant.
In second-site revertants, the mutation is at a different site in the DNA. Second-site mutations can restore a wild-type phenotype if they function as suppressor mutations—mutations that compensate for the effect of the original mutation. Several classes of suppressor mutations are known. These include (1) a mutation somewhere else in the same gene that restores enzyme function, such as a second frameshift mutation near the first that restores the original reading frame (Figure 9.6); (2) a mutation in another gene that restores the function of the original mutated gene; and (3) a mutation in another gene that results in the production of an enzyme that can replace the nonfunctional one.
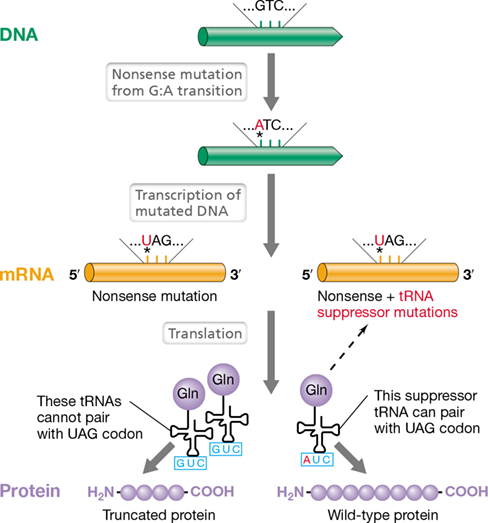
Suppressors can be best illustrated by mutations in tRNAs. Nonsense mutations can be suppressed by changing the anticodon sequence of a tRNA molecule so that it now recognizes a stop codon (Figure 9.7). Such an altered tRNA is called a suppressor tRNA and will insert the amino acid it carries at the stop codon that it now reads. Suppressor tRNA mutations exist in all three domains of life, and their activity has been observed in both Bacteria and Archaea by introducing reporter genes (Section 8.1) containing nonsense mutations. During these experiments, complete translation and activity of the corresponding reporter protein was observed despite the presence of a stop codon within the coding region of the reporter.
Figure 9.7 Suppression of nonsense mutations.

Introduction of a nonsense mutation in a gene encoding a protein results in the incorporation of a stop codon (indicated by the *) in the corresponding mRNA. This single mutation leads to the production of a truncated polypeptide. The mutation is suppressed if a second mutation occurs in the anticodon of a tRNA, a tRNA charged with glutamine in this example, which allows the mutated tRNA or suppressor tRNA to bind to the nonsense codon.
Suppressor tRNA mutations would be lethal unless a cell has more than one tRNA gene for a particular codon. One tRNA gene may then be mutated to produce a suppressor, while the other gene’s product performs the original function. Most cells have multiple genes for tRNAs, and so suppressor mutations are reasonably common, at least in microorganisms. Sometimes the amino acid inserted by the suppressor tRNA is identical to the original amino acid and the protein is fully active. In other cases, however, a different amino acid is inserted and a protein that is only partially active may be produced.
Mutation Rates
For most microorganisms, errors in DNA replication occur at a frequency of 10−6 to 10−7 per thousand bases during a single round of replication. A typical gene has about 1000 base pairs. Therefore, the frequency of a mutation in a given gene is also in the range of 10−6 to 10−7 per round of replication. For instance, in a bacterial culture having 108 cells/ml, there are likely to be a number of different mutants for any given gene in each milliliter of culture. Eukaryotes with very large genomes tend to have replication error rates about 10-fold lower than typical bacteria, whereas DNA viruses, especially those with very small genomes, may have error rates 100-fold to 1000-fold higher than those of cellular organisms. RNA viruses have even higher error rates due to less effective polymerase proofreading (Section 6.4) and the lack of RNA repair mechanisms.
Single base errors during DNA replication are more likely to lead to missense mutations than to nonsense mutations because most single base substitutions yield codons that encode other amino acids (Table 6.4). The next most frequent type of codon change caused by a single base change leads to a silent mutation. This is because for the most part alternate codons for a given amino acid differ from each other by a single base change in the “silent” third position. A given codon can be changed to any of 27 other codons by a single base substitution, and on average, about two of these will be silent mutations, one a nonsense mutation, and the rest missense mutations.
Unless a mutation can be selected for, its experimental detection is difficult, and much of the skill of the microbial geneticist requires increasing the efficiency of mutation detection. This can be done most effectively by increasing the pool of mutations. As we see in the next section, it is possible to greatly increase the mutation rate by treatment with mutagenic agents. In addition, the mutation rate may change under certain circumstances, such as when cells are placed under high-stress conditions.
Check Your Understanding
Which class of mutation, missense or nonsense, is more common, and why?
9.4 Mutagenesis
The spontaneous rate of mutation is very low, but a variety of chemical, physical, and biological agents can increase the mutation rate and are therefore said to induce mutations. These agents are called mutagens. Because Bacteria and Archaea are often exposed to mutagens, not only intentionally in the laboratory but also accidentally in their environment, we discuss some of the major categories of mutagens and their activities here.
Chemical Mutagens and Radiation
An overview of some of the major chemical mutagens and their modes of action is given in Table 9.2. Several classes of chemical mutagens exist. The nucleotide base analogs are molecules that resemble the purine and pyrimidine bases of DNA in structure but display faulty base-pairing properties (Figure 9.8). If a base analog is incorporated into DNA in place of the natural base, the DNA may replicate normally most of the time. However, DNA replication errors occur at higher frequencies at these sites due to incorrect base pairing. The result is the incorporation of a mismatched base into the new strand of DNA and thus introduction of a mutation. During subsequent segregation of this strand in cell division, the mutation is revealed.
Table 9.2 Chemical and physical mutagens and their modes of action

Figure 9.8 Nucleotide base analogs.

Structure of two common nucleotide base analogs used to induce mutations compared with the normal nucleic acid bases for which they substitute. (a) 5-Bromouracil can base-pair with guanine, causing AT to GC substitutions. (b) 2-Aminopurine can base-pair with cytosine, causing AT to GC substitutions.
Other chemical mutagens induce chemical modifications in one base or another, resulting in faulty base pairing or related changes (Table 9.2). For example, alkylating agents (chemicals that react with amino, carboxyl, and hydroxyl groups by substituting them with alkyl groups) such as nitrosoguanidine are powerful mutagens and generally induce mutations at higher frequency than base analogs. Unlike base analogs, which have an effect only when incorporated during DNA replication, alkylating agents can introduce changes even in nonreplicating DNA. Both base analogs and alkylating agents tend to induce base-pair substitutions (Section 9.2).
Another group of chemical mutagens, the acridines, are planar molecules that function as intercalating agents. These mutagens become inserted between two DNA base pairs and push them apart. Then, during replication, this abnormal conformation can trigger single base insertions or deletions. Thus, acridines typically induce frameshift rather than point mutations (Section 9.2). Ethidium bromide, which is commonly used to detect DNA in gel electrophoresis, is also an intercalating agent and therefore a mutagen.
Nonionizing and ionizing radiation are two forms of electromagnetic radiation that are highly mutagenic (Figure 9.9). Ultraviolet (UV) radiation is widely used to generate mutations because the purine and pyrimidine bases of nucleic acids absorb UV radiation strongly (the absorption maximum for DNA and RNA is at 260 nm). The primary mutagenic effect is the production of pyrimidine dimers, in which two adjacent pyrimidine bases (cytosine or thymine) on the same strand of DNA become covalently bonded to one another. This either greatly impedes DNA polymerase activity or greatly increases the probability of DNA polymerase misreading the sequence at this point. Thus the killing of cells by UV radiation is due primarily to its effect on DNA. Conversely, ionizing radiation is more powerful than UV radiation and includes short-wavelength radiation such as X-rays, cosmic rays, and gamma rays (Figure 9.9). These rays cause water and other substances to ionize, resulting in the formation of free radicals such as the hydroxyl radical (OH⋅, Section 4.16) that can damage macromolecules in the cell, including DNA. This causes double-stranded and single-stranded breaks that may lead to rearrangements or large deletions.
Figure 9.9 Wavelengths of radiation.

Ultraviolet radiation consists of wavelengths just shorter than visible light. For any electromagnetic radiation, the shorter the wavelength, the higher the energy. DNA absorbs strongly at 260 nm.
DNA Repair and the SOS System
By definition, a mutation is a heritable change in the genetic material. Therefore, if damaged DNA can be corrected before the cell divides, no mutation will occur. While cells have a variety of different DNA repair processes to correct mistakes (Section 6.4) or repair damage, some are error-prone and the repair process itself introduces the mutation. Some types of DNA damage, especially large-scale damage from highly mutagenic chemicals or large doses of radiation, may cause lesions that interfere with replication. If such lesions are not removed before replication occurs, DNA replication will stall and lethal breaks in the chromosome will result.
In Bacteria, stalled replication or major DNA damage activates the SOS repair system. The SOS system initiates a number of DNA repair processes, some of which are error-free. However, the SOS system also allows DNA repair to occur without a template, that is, with random incorporation of nucleotide precursors (deoxyribonucleotide triphosphates [dNTPs]). As might be expected, this results in many errors and hence many mutations. However, mutations induced by the SOS repair system are better than the alternative (death of the cell), as mutations can often be corrected while chromosome breaks usually cannot.
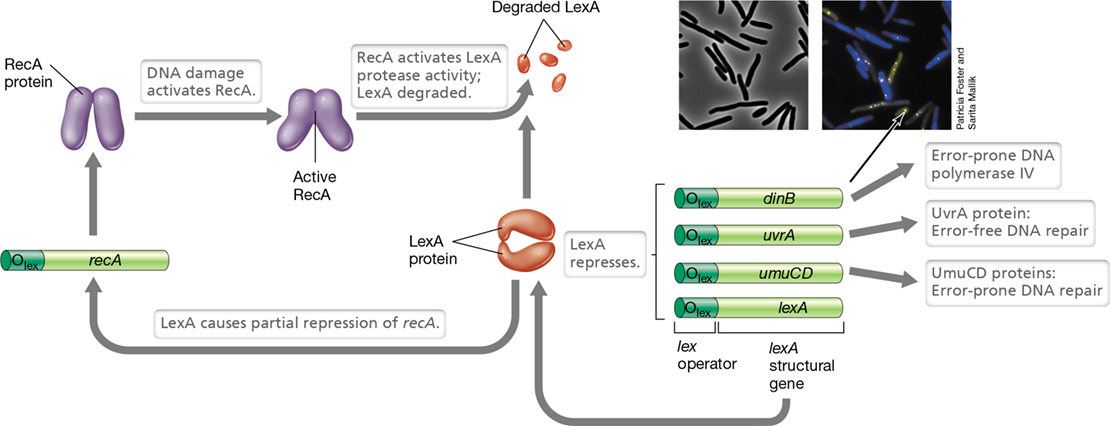
In Escherichia coli the SOS repair system controls the transcription of approximately 40 genes located throughout the chromosome that participate in DNA damage tolerance and DNA repair. Not only does the SOS system form a regulon, but the general stress response or RpoS regulon also plays a role in the repair system (Sections 7.3 and 7.9). In DNA damage tolerance, DNA lesions remain in the DNA, but are bypassed by specialized DNA polymerases that can move past DNA damage—a process called translesion synthesis. Even if no template is available to allow insertion of the correct bases, it is less dangerous to cell survival in the long run to fill the gap than to let it remain. Consequently, translesion synthesis generates many errors. In E. coli, in which the process of mutagenesis has been studied in great detail, the two error-prone repair polymerases are DNA polymerase V, an enzyme encoded by the umuCD genes, and DNA polymerase IV, encoded by dinB (Figure 9.10). Both are induced as part of the SOS repair system.
Figure 9.10 SOS response to DNA damage.

DNA damage activates RecA protein, which in turn activates the protease activity of LexA, resulting in self-cleavage. LexA normally represses the activities of RecA, the genes uvrA and umuCD that encode DNA repair functions (the UmuCD proteins are part of DNA polymerase V), and dinB, which encodes DNA polymerase IV. However, repression is not complete. Some RecA protein is produced even in the presence of LexA protein. When LexA is inactivated, DNA repair genes are highly transcribed. Inset photos: Both photos show DNA polymerase IV localization to the nucleoid during the SOS response in Escherichia coli. Cells containing a fluorescently tagged DNA polymerase IV (DinB) were treated with an antibiotic to induce DNA damage. Left: phase-contrast micrograph. Right: fluorescence micrograph showing cells stained with DAPI (blue) and DNA polymerase IV (yellow, in the nucleoid region). Expression of dinB requires not only the loss of LexA repression but also the protein RpoS, an RNA polymerase sigma factor whose synthesis is triggered by various stress responses.
The master regulators of the SOS system are the proteins LexA and RecA. LexA is a repressor that normally prevents expression of the SOS system. The RecA protein, which normally functions in genetic recombination (Section 9.5), is activated by the presence of DNA damage, in particular by the single-stranded DNA that results when replication stalls. The activated form of RecA then stimulates LexA to inactivate itself by self-cleavage. This leads to derepression of the SOS system and the coordinate expression of proteins that participate in DNA repair. Because some of the DNA repair mechanisms of the SOS system—such as DNA polymerases IV and V—are inherently error-prone, many mutations arise. However, once the original DNA damage has been repaired, the SOS regulon is repressed and further mutagenesis ceases.
Besides controlling some DNA repair systems and error-prone polymerases, the SOS system also regulates processes that contribute to horizontal gene transfer, which we discuss next. Thus, uptake and recombination of foreign DNA can also be used by Bacteria and Archaea to fix chromosomal breaks. As we shall see in Part II, not only does this form of DNA repair help with cell survival, but it also can increase cell diversity.
II Gene Transfer in *Bacteria*
**Lateral gene flow within and between species of Bacteria is highly dynamic and facilitated by at least three different mechanisms. Such genetic exchange is distinct from traditional mother-to-daughter inheritance and allows cells to rapidly acquire new characteristics and increase their competitive fitness.**
Comparative genomic analyses of closely related microbes that exhibit different phenotypes have revealed distinct genome differences. Often these idiosyncratic differences result from horizontal gene transfer, the movement of genes between cells that are not direct descendants of one another (Section 13.9). Horizontal gene transfer allows cells to quickly acquire new characteristics and fuels metabolic diversity.
Three mechanisms of genetic exchange are known in bacteria: (1) transformation, in which free DNA released from one cell is taken up by another (Section 9.6); (2) transduction, in which DNA transfer is mediated by a virus (Section 9.7); and (3) conjugation, in which DNA transfer requires cell-to-cell contact and a conjugative plasmid in the donor cell (Sections 9.8 and 9.9). While these processes were presented in Figure 9.1, they are compared and contrasted in Figure 9.11. It should be noted that DNA transfer typically occurs in only one direction, from donor to recipient.
Figure 9.11 Processes by which DNA is transferred from donor to recipient bacterial cell.

Just the initial steps in transfer are shown. Note that conjugation requires cell–cell contact, whereas transduction and transformation do not.
Before discussing the mechanisms of transfer, we consider the fate of transferred DNA. Regardless of how it was transferred, DNA that enters the cell by horizontal gene transfer faces three possible fates: (1) It may be degraded by the recipient cell’s restriction enzymes or other DNA destruction systems (Section 9.12); (2) it may replicate by itself (but only if it possesses its own origin of replication, such as a plasmid or phage genome); or (3) it may recombine with the recipient cell’s chromosome.
9.5 Genetic Recombination
Recombination is the physical exchange of DNA between genetic elements (structures that carry genetic information). Here we focus on homologous recombination, a process that results in genetic exchange between homologous DNA sequences from two different sources. Homologous DNA sequences are those that have nearly the same sequence; therefore, bases can pair over an extended length of the two DNA molecules to facilitate exchange. This type of recombination is often essential to a cell’s ability to retain DNA following genetic exchange and is behind the well-known phenomenon of “crossing over” in the genetics of eukaryotes.
Molecular Events in Homologous Recombination
The RecA protein, previously mentioned in regard to the SOS repair system (Section 9.4 and Figure 9.10), is the key to homologous recombination. RecA is essential in nearly every homologous recombination pathway. RecA-like proteins have been identified in all Bacteria examined, as well as in the Archaea and most Eukarya.
A molecular mechanism for homologous recombination between two DNA molecules is shown in Figure 9.12. An enzyme that cuts DNA in the middle of a strand, called an endonuclease, begins the process by nicking one strand of the donor DNA molecule. This nicked strand is separated from the other strand by proteins with helicase activity; the resulting single-stranded segment binds single-strand binding protein (Section 6.3) and then RecA. This results in a complex that promotes base pairing with the complementary sequence in the recipient DNA molecule. Base pairing, in turn, displaces the other strand of the recipient DNA molecule (Figure 9.12) and is appropriately called strand invasion.
Figure 9.12 Homologous recombination.
Homologous DNA molecules pair and exchange DNA segments. The mechanism requires breakage and reunion of paired segments, as indicated by red arrows. Two of the participating proteins, single-strand binding (SSB) protein and the RecA protein, are shown (other participating proteins are not shown). Resolution occurs by cutting and rejoining the cross-linked DNA molecules. Note that there are two possible outcomes, patches or splices, depending on where strands are cut during the resolution process.
The base pairing of one strand from each of the two DNA molecules over long stretches generates recombination intermediates containing long heteroduplex regions, where each strand has originated from a different chromosome. After DNA strands at the branch or crossover regions are joined, the linked molecules are then resolved (separated) by enzymes that cut and rejoin the previously unbroken strands of both original DNA molecules. Depending on the orientation of the junction during resolution, two types of products—referred to as “patches” or “splices”—are formed that differ in the conformation of the heteroduplex regions remaining after resolution (Figure 9.12).
Effect of Homologous Recombination on Genotype
For homologous recombination to generate new genotypes, the two homologous sequences must be related but genetically distinct. This is obviously the case in a diploid eukaryotic cell, which has two sets of chromosomes, one from each parent. However, in bacteria, genetically distinct but homologous DNA molecules are brought together in different ways. Genetic recombination in bacteria occurs after fragments of homologous DNA from a donor chromosome are transferred to a recipient cell by transformation, transduction, or conjugation. It is only after the transfer event, when the DNA fragment from the donor is in the recipient cell, that homologous recombination occurs.
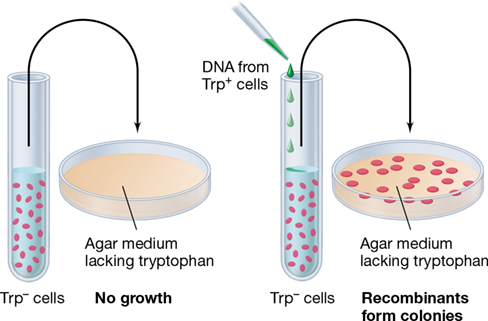
For physical exchange of DNA segments to be detected, the cells resulting from recombination must be phenotypically different from both parents (Figure 9.13). Genetic crosses in bacteria usually depend on using recipient strains that lack some selectable character that the recombinants will gain. For instance, the recipient may be unable to grow on a particular medium or may exhibit a specific phenotype due to a point mutation, while the genetic recombinants can grow on the particular medium or exhibit a phenotype different from the recipient (Figures 9.2 and 9.13). Various kinds of selectable markers, such as drug resistance and nutritional requirements, were discussed in Section 9.1. The exceedingly great sensitivity of the selection process allows even a few recombinant cells to be detected in a large population of nonrecombinant cells, and thus selection is an important tool for the microbial geneticist.
Figure 9.13 Using a selective medium to detect rare genetic recombinants.

On the selective medium, only the rare recombinants form colonies even though a very large population of bacteria is plated. Procedures such as this, which offer high resolution for genetic analyses, can ordinarily be used only with microorganisms. The type of genetic exchange being illustrated is transformation, but a similar outcome could result from any of the other forms of horizontal gene transfer (Figure 9.1).
Complementation
In all three methods of bacterial gene transfer, only a portion of the donor chromosome enters the recipient cell and thus transfer is just the first step; unless recombination takes place with the recipient chromosome, the donor DNA will be lost because it cannot replicate independently in the recipient. Nonetheless, it is possible to stably maintain a state of partial diploidy for use in bacterial genetic analysis. A bacterial strain that carries two copies of any particular chromosomal segment is known as a partial diploid, or merodiploid. In general, one copy is present on the chromosome itself and the second copy on another genetic element, such as a plasmid or a bacteriophage.
Consequently, if the chromosomal copy of a gene is defective due to a mutation, it is possible to supply a functional (wild-type) copy of the gene on a plasmid or bacteriophage. For example, if one of the genes for biosynthesis of the amino acid tryptophan has a mutation resulting in a nonfunctional enzyme, this will yield a Trp− phenotype (the mutant strain will be a tryptophan auxotroph and must be supplied with tryptophan for growth). However, if a copy of the wild-type gene is introduced into the same cell on a plasmid or viral genome, this gene will encode the necessary protein and, assuming the gene is transcribed and translated, will restore the wild-type phenotype. This process is called complementation because the wild-type gene is said to complement the mutation, in this case converting the Trp− cell into Trp+ (Figure 9.13).
Check Your Understanding
Which protein, found in virtually all cells, facilitates the pairing required for homologous recombination?
Explain the fate of transferred chromosomal DNA if recombination does not occur in the recipient cell.
What is a merodiploid, and what is genetic complementation?
9.6 Transformation
Transformation is a genetic transfer process by which free DNA is incorporated into a recipient cell and brings about genetic change. When would a cell encounter free DNA? Because the DNA in bacterial and archaeal cells is present as a large single molecule, chromosomal DNA pours out during cell lysis. Cells that are naturally transformable, which include certain species of both gram-negative and gram-positive Bacteria and also some species of Archaea (Section 9.10), can take up pieces of this free DNA from their lysed neighbors (Figures 9.1 and 9.11).
Because of their extreme length, bacterial chromosomes break easily. For example, the Bacillus subtilis chromosome would be 1700 μm long if linearized. However, even gentle DNA extraction methods result in the B. subtilis 4.2-megabase-pair chromosome being broken into fragments of about 10 kilobase pairs each. Because an average gene contains about 1000 nucleotides, each of the fragments of B. subtilis DNA contains about ten genes. This is a typical transformable size. A single cell typically incorporates only one or at most a few DNA fragments, so only a small proportion of the genes of one cell can be transferred to another in a single transformation event.
Competence in Transformation
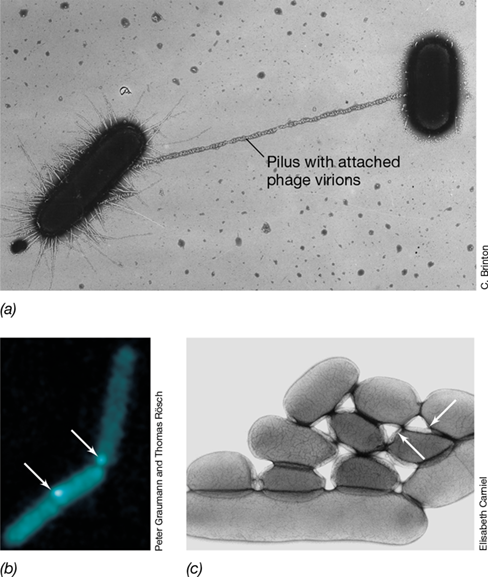
How does a cell take up DNA? Recall from our Chapter 2 discussion that cell membranes are selective barriers. DNA does not freely cross this barrier; thus a cell that is able to take up DNA and be transformed is said to be competent. Figure 9.14 illustrates Vibrio cholerae, the causative agent of cholera and a model gram-negative bacterium for natural competence, taking up DNA. In its natural marine environment, V. cholerae aggressively competes for nutrients by using its type VI secretion system to inject toxin molecules into neighboring cells (Section 6.13). These toxin molecules ultimately kill competitor cells. Not only do the V. cholerae cells obtain nutrients from lysed neighboring cells, but they are also able to scavenge the DNA released from their victims (Figure 9.14).
Figure 9.14 ***Vibrio cholerae*** killing neighboring prey cells and scavenging DNA.

Time-course images of V. cholerae type VI secretion system proteins labeled with a green fluorescent protein (GFP) and corresponding DNA uptake proteins labeled with a red fluorescent protein. V. cholerae cells expressing both protein types appear red with green “crossbows” inside, while competing cells lacking a type VI secretion system appear as solid green cells. At time 0, white arrows indicate prey cells that have been killed. These cells circularize upon death. After 30 minutes, DNA uptake proteins localize for effective uptake of DNA from killed cells (circled). Images modified from Borgeaud, S., Metzger, L.C., Scrignari, T., and Blokesch, M. 2015. Science 347: 63–67.
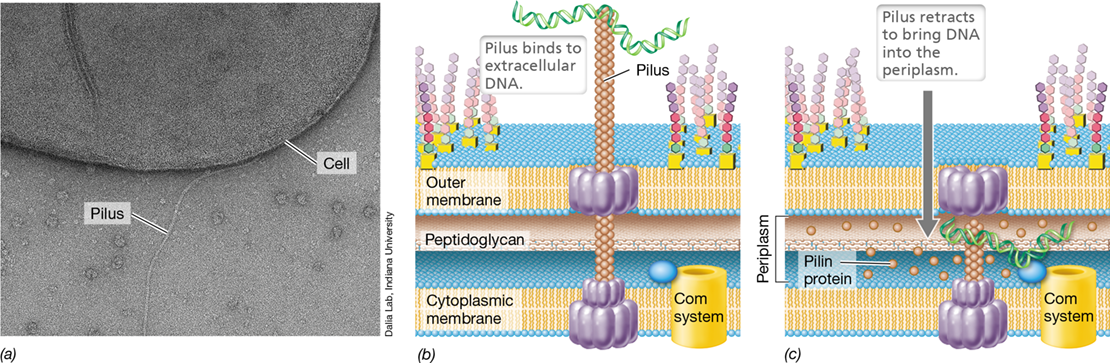
The capacity to take up free DNA is genetically determined. In Bacteria such as Vibrio, Neisseria, Acinetobacter, Thermus, Streptococcus, and Bacillus, competence is directly linked to pili (Figure 9.15). Pili are versatile multiprotein structures that not only play roles in twitching, gliding motility, biofilm formation, and pathogenesis (Sections 2.6 and 2.10), but are also key to binding and taking up DNA in naturally competent cells. In gram-negative Bacteria, pili cross the hydrophobic outer membrane through a specific channel, and protein subunits within the pilus recognize and bind to extracellular DNA (Figure 9.15b). After binding to DNA, V. cholerae pili pull the DNA through the outer membrane into the periplasm through a process called pilus retraction (see MicrobiologyNow in the chapter opener). This retraction of the pilus is the result of depolymerization of individual pilin proteins that make up the pilus structure (Figure 9.15c). In gram-positive cells, pili or homologs of type II secretion systems (Section 6.13) are used to bind DNA and bring it through the thick cell wall layer. Once in the periplasm of gram-negative cells or through the cell wall in gram-positive cells, the DNA binds to proteins within the competence system associated with the cytoplasmic membrane. These competence-specific proteins include a membrane-associated DNA-binding protein and various nucleases (Figure 9.16).
Figure 9.15 *Vibrio* pilus and DNA uptake.

(a) Transmission electron micrograph showing Vibrio cholerae type IV pilus. (b) The pilus binds extracellular DNA, and (c) pilus retraction brings extracellular DNA through the outer membrane to the competence (Com) system associated with the cytoplasmic membrane. See the photomicrograph of a V. cholerae cell using its pilus to retrieve DNA on page 261.
Figure 9.16 General mechanism of transformation.

To begin, double-stranded DNA is bound by a membrane-bound DNA-binding protein. Passage of one of the two strands of DNA into the cell is followed by nuclease activity that degrades the complementary strand. Once the single strand of DNA in the cell is bound by specific proteins, recombination with homologous regions of the bacterial chromosome is mediated by the RecA protein with the final product being a transformed cell.
As noted earlier, the sizes of the transforming fragments are much smaller than that of the whole genome, and the fragments are further degraded during the uptake process by competence proteins (Figure 9.16). As these fragments are taken up into the cytoplasm, they are often converted into single-stranded pieces of DNA, with the complementary strand being degraded. After entry into the cytoplasm, a competence-specific protein binds the single-stranded donor DNA. This protects the DNA from nuclease attack until it reaches the recipient’s chromosome, where the RecA protein takes over. If the transforming DNA shares sequence homology with that of the chromosome, then it is integrated into the genome of the recipient by recombination (Figures 9.12 and 9.16).
The preceding applies only to small pieces of linear DNA. Many naturally transformable Bacteria are transformed only poorly by plasmid DNA because the plasmid must remain double-stranded and circular in order to replicate. The success rate of natural plasmid DNA transformation can be enhanced through delivery from a donor to a recipient cell through extracellular vesicles (Figure 9.1), which we will consider in more detail in Archaea in Section 9.10.
Regulation of Competence
Competence in most naturally transformable bacteria is regulated. One pathway of natural competence in Bacillus subtilis—an easily transformed species—is regulated by quorum sensing, a regulatory system that responds to cell density (Section 7.7). Cells produce and excrete a small peptide during growth, and the accumulation of this peptide to high concentrations induces the cells to become competent. But not all cells in a population become competent. In Bacillus, roughly 20% of the cells become competent and stay that way for several hours. By contrast, in Streptococcus, 100% of the cells can become competent, but only for a brief period during the growth cycle.
Multiple layers of regulation control natural competence in other bacteria. Vibrio cholerae is naturally found in marine and freshwater environments associated with crustacean exoskeletons, which are composed of chitin. Competence in V. cholerae is controlled not only by quorum sensing but also by chitin sensing and catabolite repression (Section 7.8). V. cholerae can catabolize chitin (a polymer of N-acetylglucosamine and an abundant nutrient in the marine environment), and as cells aggregate on a chitin surface, they are in close proximity to one another and more likely to successfully exchange DNA (Figure 9.17).
Figure 9.17 Regulation of natural competence in *Vibrio cholerae*.

V. cholerae cells with fluorescent reporter genes linked to the promoters of competence genes were grown in the presence of chitin beads. White dashed line indicates edge of bead surface. (a) Cells with the pilA (pilus protein) promoter linked to a green fluorescent protein (GFP). (b) Cells with the comEA (DNA uptake proteins) promoter linked to a red fluorescent protein. (c) Merged image of parts a and b illustrating expression of competence genes in cells associated with the chitin bead. Cells of V. cholerae are about 0.5 μm wide and 1.5 μm long. Adapted from Lo Scrudato, M., and M. Blokesch. 2012. PLoS Genetics 8(6): e1002778. See Figure 9.14 for a different view of transformation in V. cholerae.
High-efficiency, natural transformation is rare among Bacteria. For example, Acinetobacter, Bacillus, Streptococcus, Haemophilus, Neisseria, and Thermus are naturally competent and easy to transform. This natural competence provides a nutritional advantage, as free DNA is rich in carbon, nitrogen, and phosphorus. By contrast, many Bacteria are poorly transformed, if at all, under natural conditions. For example, Escherichia coli and many other gram-negative bacteria fall into this category. However, if cells of E. coli are treated with high concentrations of Ca2+ and then chilled, they become adequately competent. Cells treated in this manner take up double-stranded DNA, and therefore transformation of E. coli by plasmid DNA can be relatively efficient. This is important because getting DNA into E. coli—the workhorse of genetic engineering—is critical for biotechnology, as we will see in Chapter 12.
Electroporation is a physical technique that is used to get DNA into organisms that are difficult or impossible to transform, especially cells that contain thick cell walls. In electroporation, cells are mixed with DNA and then exposed to brief, high-voltage electrical pulses. This makes the cell envelope permeable and allows entry of the DNA. Electroporation works for getting free DNA into most types of cells, including E. coli, most other Bacteria, some species of Archaea, and even yeast and certain plant cells.
Check Your Understanding
During transformation a cell usually incorporates only one or a few fragments of DNA. Explain.
In genetic transformation, what is meant by the word competence?
During natural competence, what type of cellular structure is used to bind DNA and take it to the cytoplasmic membrane?
9.7 Transduction
In transduction, a bacterial virus (bacteriophage, Chapter 5) transfers DNA from one cell to another. Viruses can transfer host genes in two ways. In the first, called generalized transduction, DNA derived from virtually any portion of the host genome is packaged inside the mature virion in place of the virus genome. In the second, called specialized transduction, DNA from a specific region of the host chromosome is integrated directly into the virus genome—usually replacing some of the virus genes. This occurs only with certain temperate viruses such as phage lambda (Section 5.6).
In generalized transduction, the bacterial donor genes cannot replicate independently and are not part of a viral genome. Thus, unless the donor genes recombine with the recipient bacterial chromosome, they will be lost. In specialized transduction, homologous recombination may also occur. However, since the donor bacterial DNA is actually a part of a temperate phage genome, it may be integrated into the host chromosome during lysogeny (Section 5.6). This can lead to an event called phage conversion (see later).
Transduction occurs in a variety of Bacteria, including the genera Desulfovibrio, Escherichia, Pseudomonas, Rhodococcus, Rhodobacter, Salmonella, Staphylococcus, and Xanthobacter, as well as Methanothermobacter thermautotrophicus, a species of Archaea. Not all phages can transduce and not all bacteria are transducible. However, considering the enormous genetic diversity of viruses (Chapter 11) and the fact that bacteriophages are estimated to outnumber prokaryotic cells in nature by 10-fold, transduction likely plays an important role in gene transfer in the environment for many species of Bacteria and Archaea. Some examples of genes transferred by transducing bacteriophages include multiple-antibiotic-resistance genes among strains of Salmonella enterica (typhimurium), Shiga-like toxin genes in Escherichia coli, virulence factors in Vibrio cholerae, and genes encoding photosynthetic proteins in cyanobacteria (Section 20.13).
Generalized Transduction
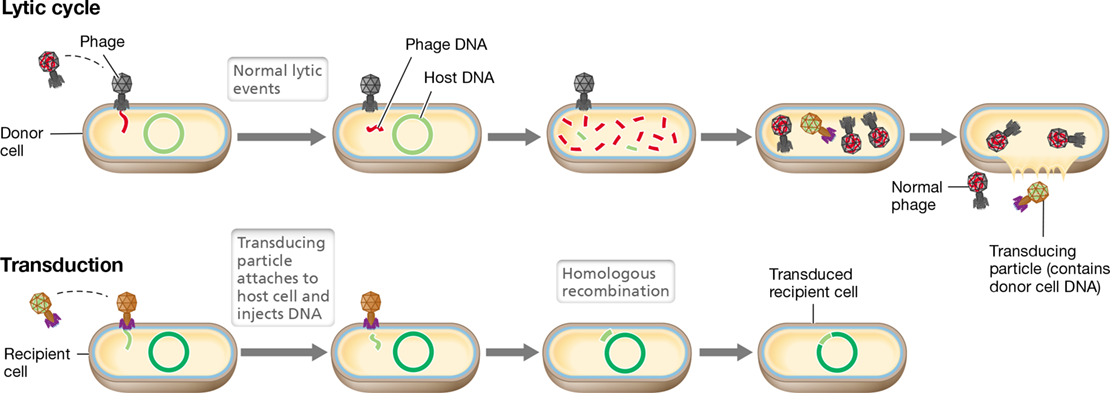
In generalized transduction, virtually any gene on the donor chromosome can be transferred to the recipient, forming a transductant. Generalized transduction was first discovered and extensively studied in the bacterium S. enterica with phage P22 and has also been studied with phage P1 in E. coli. The mechanism of transduction is shown in Figure 9.18. When a bacterial cell is infected with a transducing phage, the lytic cycle may occur. However, during lytic infection, the enzymes responsible for packaging viral DNA into the bacteriophage sometimes package host DNA accidentally. The result is called a transducing particle. These cannot lead to a viral lytic infection because they contain no viral DNA, and are therefore said to be defective.
Figure 9.18 Generalized transduction.
Note that “normal” virions contain phage genes, whereas a transducing particle contains host genes.
Mastering Microbiology
Art Activity: Figure 9.18 Generalized transduction
Upon lysis of the cell, transducing particles are released along with normal virions that contain the virus genome. When this lysate is used to infect a population of recipient cells, most of the cells are infected with a normal (lytic) virus. However, a small proportion of the population receives transducing particles that inject the DNA they packaged from the previous host bacterium. Although this DNA cannot replicate, it can recombine with the DNA (Section 9.5) of the new host (Figure 9.19). Because only a small proportion of the particles in the lysate are defective, and each of these contains only a small fragment of donor DNA, the probability of a given transducing particle containing a particular gene is quite low. Typically, only about 1 cell in 106 to 108 cells is transduced for any given gene.
Figure 9.19 Visualization of generalized transduction.

Citrobacter freundii cells were mixed with P1 bacteriophage carrying the β-lactamase (bla) gene for 10 minutes. (a) Fluorescence micrograph showing cells by DAPI staining. (b) Detection of a single C. freundii transductant containing the bla gene recombined into the genome as visualized by fluorescence in situ hybridization (FISH, Section 19.5). Arrows indicate the cell that is transduced.
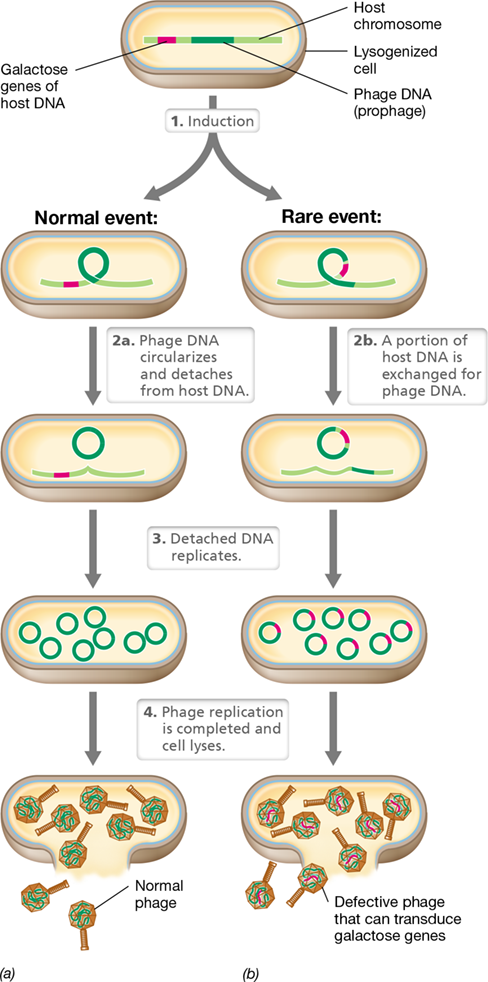
Lysogeny and Specialized Transduction
Generalized transduction allows the transfer of any gene from one bacterium to another, but at a low frequency. In contrast, specialized transduction allows extremely efficient transfer but is selective and transfers only a small region of the bacterial chromosome. In the first case of specialized transduction to be discovered, genes for galactose catabolism were transduced by the temperate phage lambda of E. coli.
Mastering Microbiology
Art Activity: Figure 9.20 Specialized transduction
When lambda lysogenizes a host cell, the phage genome is integrated into the E. coli chromosome at a specific site (Section 5.6 and Section 11.4). This site is next to the cluster of genes that encode the enzymes for galactose utilization. After insertion, viral DNA replication is under control of the bacterial host chromosome. Upon induction, the viral DNA separates from the host DNA by a process that is the reverse of integration (Figure 9.20). Usually the lambda DNA is excised precisely, but occasionally the phage genome is excised incorrectly. Some of the adjacent bacterial genes to one side of the prophage (for example, the galactose operon) are excised along with phage DNA. At the same time, some phage genes are left behind (Figure 9.20b). This transducing particle can subsequently transfer genes for galactose utilization to a recipient cell. This transfer can only be detected if a galactose-negative (Gal−) strain is infected with such a transducing particle and Gal+ transductants are selected.
Figure 9.20 Specialized transduction.

In an Escherichia coli cell containing a lambda prophage, (a) normal lytic events and (b) the production of particles transducing the genes for galactose utilization. Only a short region of the circular host chromosome is shown in the figure.
For a lambda virion to be infectious, there is a limit to the amount of phage DNA that can be replaced with host DNA. Sufficient phage DNA must be retained to encode the phage protein coat and other phage proteins needed for lysis and lysogeny. However, if a helper phage is used together with a defective phage in a mixed infection, then far fewer phage-specific genes are needed in the defective phage. Only the att (attachment) region, the cos site (cohesive ends, for packaging), and the replication origin of the lambda genome (Figure 11.11) are necessary.
Phage Conversion
Alteration of the phenotype of a host cell by lysogenization is called phage conversion. When a normal (that is, nondefective) temperate phage lysogenizes a cell and becomes a prophage, the cell becomes immune to further infection by the same type of phage. Such immunity may itself be regarded as a change in phenotype. However, other phenotypic changes unrelated to phage immunity are often observed in phage conversion of lysogenized cells due to foreign genes present in the phage genome.
Overall, the genes responsible for the phage conversion are an integral part of the phage genome and hence are automatically transferred to the cell upon phage infection and establishment of the lysogenic state. This lysogenic state likely carries strong selective value for the host cell because it confers resistance to infection by viruses of the same type. It has also been found that many bacteria isolated from nature are natural lysogens and that their corresponding prophage encode an abundance of transposons (Section 9.11 and Section 13.9). Thus it is likely that phage conversion contributes to the formation of new genes that are essential for survival of many host cells in nature.
Cases of phage conversion that promote bacterial virulence have been especially well studied. Vibrio cholerae, which causes the disease cholera (Section 33.3), is a fascinating example of how phage conversion can result in a nonpathogenic environmental strain evolving into a pathogen. CTXφ is a lysogenic bacteriophage found integrated into the chromosome of pathogenic V. cholerae strains. Besides the normal genes necessary to maintain the lysogenic state, the CTXφ prophage also encodes the cholera toxin that triggers the diarrheal disease. During infection, CTXφ virions specifically bind to the toxin co-regulated pilus (TCP) used by V. cholerae to attach to host intestinal cells (Section 33.3). Interestingly, this TCP appears to be encoded by a separate prophage. Strains of Vibrio that are not lysogenized by CTXφ or TCP are nonpathogenic, thus highlighting the power of phage conversion in enhancing genetic fitness and pathogen evolution. In later chapters we will see many other human infectious diseases in which phage conversion plays a critical role, including in such distinct diseases as whooping cough and botulism.
Gene Transfer Agents
DNA can also be transferred between prokaryotic cells by defective bacteriophages. These so-called gene transfer agents (GTAs) are the result of prokaryotic cells hijacking defective viruses and using them specifically for DNA exchange (Figure 9.21). GTAs resemble tiny tailed bacteriophages and contain random small pieces of host DNA. They are not considered true viruses because they do not contain genes encoding their own production and do not produce characteristic viral plaques. Instead, the genes encoding GTAs lie within the genome of the cell that produces them, while other regions of the genome are packaged within the agents.
Figure 9.21 Gene transfer agents.

Inset: Transmission electron micrograph of a gene transfer agent (GTA) isolated from Rhodobacter capsulatus. Visualization of a subset of R. capsulatus cells producing and releasing GTAs during stationary phase using a red fluorescent reporter gene linked to the promoter of an R. capsulatus gene essential for GTA production. Note that only a subset of the total population produces GTA.
GTAs have been isolated from a wide range of Bacteria including the sulfate-reducing bacterium Desulfovibrio desulfuricans, a variety of phototrophic purple bacteria and other Alphaproteobacteria, and also from certain methanogenic Archaea. GTAs seem to be particularly widely produced by marine Bacteria, especially purple bacteria such as Roseovarius. GTA synthesis is regulated, and in the purple bacterium Rhodobacter capsulatus, GTA synthesis is triggered by the cell’s sensing of the stationary growth phase and nutrient fluctuations (Figure 9.21). These conditions lead to only a small subpopulation of R. capsulatus cells lysing and releasing the GTAs. However, the same conditions that trigger GTA synthesis in some cells also lead to increased expression of the genes encoding products necessary for the uptake of GTAs. In R. capsulatus, stationary phase and nutrient fluctuations lead to the increased expression of genes encoding the capsular polysaccharide (Figure 2.16) receptor that GTAs specifically bind to and natural competence proteins needed for DNA uptake. This suggests that GTAs may have evolved as a mechanism for a subpopulation of cells to sacrifice themselves in order to disperse genes into the environment in a protected form to benefit the rest of their population.
While bacteriophages are considered the most abundant microbes on Earth, the percentage of these that are actually GTAs instead of viruses is unknown, but could be significant. The fact that GTAs are produced by so many different species points to GTAs as major vehicles for gene flow between prokaryotic cells in nature. This could be especially true of open-ocean microbial communities, where constant low nutrient levels might trigger GTA production as a means for cells to scavenge each other’s genes in an effort to improve fitness and survival of the overall population.
Check Your Understanding
How does a transducing particle differ from an infectious bacteriophage?
What is the major difference between generalized transduction and transformation?
Why is phage conversion considered beneficial to host cells?
9.8 Conjugation
Conjugation is a form of horizontal gene transfer that requires cell-to-cell contact (mating) and occurs in many species of gram-negative and gram-positive bacteria. Conjugation is a plasmid-encoded mechanism that can mediate DNA transfer between closely related cells or between more distantly related cells (for example, between cells of different genera). Conjugative plasmids use conjugation to transfer copies of the genes that encode themselves and other genes they may contain, such as those that encode antibiotic resistance, to new host cells.
Mastering Microbiology
Activities: Microbiology Animation: Conjugation: F Factor
The process of conjugation requires a donor cell, which contains the conjugative plasmid, and a recipient cell, which does not. In addition, genetic elements that cannot transfer themselves can sometimes be mobilized or transferred during conjugation. These other genetic elements can be other plasmids or the host chromosome itself. Indeed, conjugation was discovered because the F plasmid of Escherichia coli can mobilize the host chromosome (see Figure 9.27). Transfer mechanisms may differ depending on the participating plasmid, but most plasmids in gram-negative Bacteria employ a mechanism similar to that used by the F plasmid.
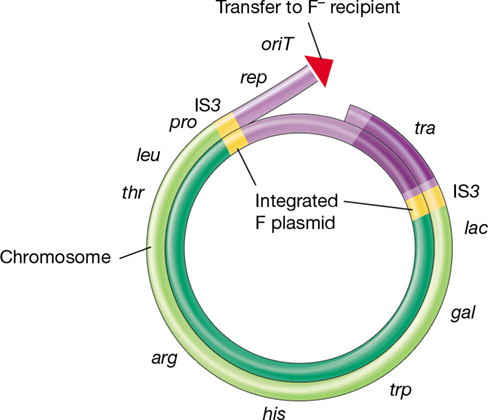
F Plasmid
The F plasmid (F stands for “fertility”) is a circular DNA molecule of 99,159 bp. Figure 9.22 shows a genetic map of the F plasmid. One region of the plasmid contains genes that regulate DNA replication. It also contains a number of transposable elements (Section 9.11) that allow the plasmid to integrate into the host chromosome. In addition, the F plasmid has a large region of DNA, the tra region, containing genes that encode transfer functions. Many genes in the tra region participate in mating pair formation, and most of these have to do with the synthesis of the conjugative pilus (Section 2.6) and a type IV secretion system (Section 6.13) to transfer the DNA. Only donor cells produce these pili. Different conjugative plasmids may have slightly different tra regions, and the pili may vary somewhat in structure. The F plasmid and its relatives encode F pili.
Figure 9.22 Genetic map of the F (fertility) plasmid of *Escherichia coli*.

The numbers on the interior mark the size of the plasmid in kilobase pairs (the exact size is 99,159 bp). The region in dark green at the bottom of the map contains genes primarily responsible for the replication and segregation of the F plasmid. The origin of vegetative replication is oriV. The light green tra region contains the genes needed for conjugative transfer. The origin of transfer during conjugation is oriT. The red arrowhead at oriT indicates the leading edge and direction of the transfer, and the black arrow is alongside the first part of the plasmid to be transferred behind oriT. The tra region is transferred last. Insertion sequences are shown in yellow. These may recombine with identical elements on the bacterial chromosome, which leads to integration and the formation of different Hfr strains (Section 9.9). Inset: Atomic force microscopy visualization of supercoiled plasmid DNA. Modified from Witz, G. and Stasiak, A. 2010. Nucleic Acids Res. 38 : 2119.
Pili allow specific pairing to take place between the donor and recipient cells. All conjugation in gram-negative Bacteria is thought to depend on cell pairing brought about by pili. The pilus makes specific contact with a receptor on the recipient cell surface and then is retracted by disassembling its subunits. This pulls the two cells together (**Figure 9.23*a***). Following this process, donor and recipient cells remain in contact by binding coupling proteins located in the outer membrane of each cell (Figure 9.23b). DNA is then transferred from donor to recipient cell through this conjugation junction (Figure 9.23c).
Figure 9.23 Visualization of conjugation.

(a) Formation of a mating pair. Direct contact between two conjugating Escherichia coli cells is first made via a pilus. The cells are then drawn together to form a mating pair by retraction of the pilus, which is achieved by depolymerization. Certain small phages (F-specific bacteriophages) use the conjugative pilus as a receptor and can be seen here attached to the pilus. (b) Coupling proteins near the cell membrane (white arrows). These Bacillus subtilis cells contain the conjugative plasmid pLS20 encoding the VirD coupling protein linked to a fluorescent reporter gene. (c) Conjugation junctions. Negatively stained transmission electron micrograph of conjugation bridges between cells of Yersinia pseudotuberculosis. Arrows indicate connection sites. Adapted from Lesic, B., M. Zouine, M. Ducos-Galand, C. Huon, M-L. Rosso, M-C. Prévost, D. Mazel, and E. Carniel. 2012 PLoS Genet. 8(3): e1002529.
Mechanism of DNA Transfer during Conjugation
DNA synthesis is necessary for DNA transfer by conjugation—the donor cell does not lose the genes it transfers but instead makes a copy as the DNA is transferred. In this case, DNA is synthesized not by normal bidirectional replication (Section 6.4) but by rolling circle replication, a mechanism also used by some DNA viruses (Section 11.3) and shown in Figure 9.24. DNA transfer is triggered by cell-to-cell contact, at which time one strand of the circular plasmid DNA is nicked and transferred to the recipient. The nicking enzyme required to initiate the process, TraI, is encoded by the tra operon of the F plasmid. TraI also has helicase activity and thus unwinds the DNA strand to be transferred. As the transfer occurs, DNA synthesis by the rolling circle mechanism replaces the transferred strand in the donor, while a complementary DNA strand is being made in the recipient. Therefore, at the end of the process, both donor and recipient possess complete plasmids. For transfer of the F plasmid, if an F-containing donor cell, which is designated F+, mates with a recipient cell lacking the plasmid (F−), the result is two F+ cells (Figure 9.24).
Figure 9.24 Transfer of plasmid DNA by conjugation.

(a) Transfer of the F plasmid converts an F− recipient cell into an F+ cell. Note the mechanism of rolling circle replication in steps 4 and 5. (b) Details of the replication and transfer process. Note the large number of proteins needed for successful DNA transfer. (c) Visualization of DNA transfer by conjugation in Bacillus subtilis using fluorescence microscopy. The donor cell constitutively expresses a red fluorescent protein, while the recipient cells fluoresce green due to green fluorescent protein (GFP) fused to LacI. The DNA transferred from the donor contains a lacO operator region that binds LacI-GFP. Arrow indicates focal point in the recipient cell where LacI-GFP is bound to the lacO region obtained from conjugation.The lac operon is discussed in Sections 7.3 and 7.8.
Transfer of plasmid DNA is efficient and rapid; under favorable conditions virtually every recipient cell that pairs with a donor acquires a plasmid. Transfer of the F plasmid (approximately 100 kbp) takes about 5 min. If plasmid genes can be expressed in the recipient, the recipient itself becomes a donor and can transfer the plasmid to other recipients. In this fashion, conjugative plasmids can spread rapidly among bacterial populations, behaving much like infectious agents. This is of major ecological significance because conjugative plasmids have been found in many Bacteria and some Archaea (Section 9.10), and a few plasmid-containing cells introduced into a population of potential recipients can convert the entire population into plasmid-bearing (and thus donating) cells in a short time.
Check Your Understanding
In conjugation, how are donor and recipient cells brought into contact with each other?
Explain how rolling circle DNA replication allows both donor and recipient to end up with a complete copy of a plasmid transferred by conjugation.
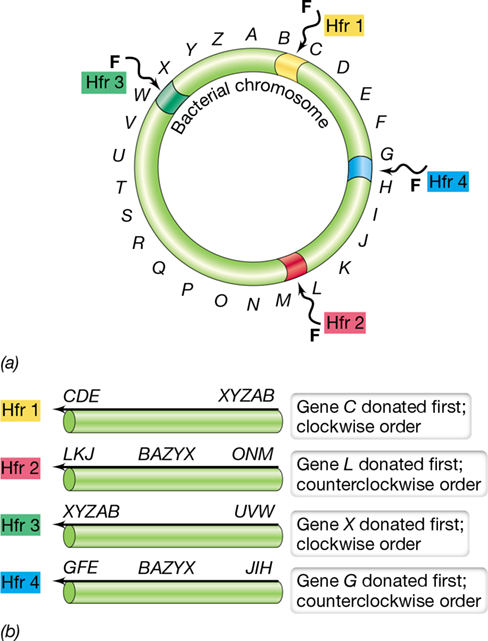
9.9 The Formation of Hfr Strains and Chromosome Mobilization
9.9 The Formation of Hfr Strains and Chromosome Mobilization
9.9 The Formation of Hfr Strains and Chromosome Mobilization
In addition to plasmid genes, chromosomal genes can be transferred by plasmid-mediated conjugation. As mentioned above, the F plasmid of Escherichia coli can, under certain circumstances, mobilize the chromosome for transfer during cell-to-cell contact. The F plasmid is actually an episome, a plasmid that can integrate into the host chromosome. When the F plasmid is integrated, chromosomal genes can be transferred along with the plasmid. Following genetic recombination between donor and recipient DNA, horizontal transfer of chromosomal genes by this mechanism can be extensive.
Cells possessing a nonintegrated F plasmid are called F+, whereas those with an F plasmid integrated into the chromosome are called Hfr cells (for high frequency of recombination). This term refers to the high rates of genetic recombination between genes on the donor (Hfr) and recipient (F −) chromosomes. Both F+ and Hfr cells are donors, but unlike conjugation between an F+ and an F −, conjugation between an Hfr donor and an F − leads to transfer of genes from the host chromosome. This is because the chromosome and plasmid now form a single molecule of DNA. Consequently, when rolling circle replication is initiated by the F plasmid, replication continues on into the chromosome. Thus, the chromosome is also replicated and parts of it get transferred. Hence, integration of a conjugative plasmid provides the mechanism for mobilizing a cell’s genome.
Mastering Microbiology
Activities: Microbiology Animation: Conjugation: Hfr Conjugation
Overall, the presence of the F plasmid results in three distinct changes in a cell: (1) the ability to synthesize the F pilus (Figure 9.23a), (2) the mobilization of DNA for transfer to another cell, and (3) the alteration of surface receptors so the cell can no longer be a recipient in conjugation and is therefore unable to take up a second copy of the F plasmid or any genetically related plasmids.
Integration of F Plasmid and Chromosome Mobilization
The F plasmid and the chromosome of E. coli both carry several copies of mobile genetic elements called insertion sequences (ISs; Section 9.11). These provide regions of sequence homology between chromosomal and F plasmid DNA. Consequently, homologous recombination between an IS on the F plasmid and a corresponding IS on the chromosome results in integration of the F plasmid into the host chromosome (Figure 9.25). Once integrated, the plasmid no longer replicates independently, but the tra operon still functions normally and the Hfr strain synthesizes pili (Figure 9.23). When a recipient cell is encountered, conjugation is triggered just as in an F+ cell, and DNA transfer is initiated at the oriT (origin of transfer) site. However, because the plasmid is now part of the chromosome, after part of the plasmid DNA is transferred, chromosomal genes begin to be transferred (Figure 9.26). As in the case of conjugation with just the F plasmid itself (Figure 9.24), chromosomal transfer also requires DNA replication.
Figure 9.25 The formation of an Hfr strain.

Integration of the F plasmid into the chromosome may occur at a variety of specific sites where IS elements are located. The example shown here is an IS3 located between the chromosomal genes pro and lac. Some of the genes on the F plasmid are shown. The arrow indicates the origin of transfer, oriT, with the arrow as the leading end. Thus, in this Hfr, pro would be the first chromosomal gene to be transferred and lac would be among the last.
Figure 9.26 Transfer of chromosomal genes by an Hfr strain.

The Hfr chromosome breaks at the origin of transfer within the integrated F plasmid. The transfer of DNA to the recipient begins at this point. DNA replicates during transfer as for a free F plasmid (Figure 9.24). This figure is not to scale; the inserted F plasmid is actually less than 3% of the size of the Escherichia coli chromosome.
Because the DNA strand usually breaks during transfer, only part of the donor chromosome is typically transferred. Consequently, the recipient does not become Hfr (or F+) because only part of the integrated F plasmid is transferred (Figure 9.27). However, after transfer, the Hfr strain remains genetically Hfr because it retains a copy of the integrated F plasmid. Following recombination, the recipient cell may express a new phenotype due to the incorporation of donor genes, but genetically it remains an F − cell. Because a partial chromosome cannot replicate, for incoming donor DNA to survive, it must recombine with the recipient chromosome. As in transformation and transduction, genetic recombination between donor and recipient genes requires homologous recombination (Figure 9.12) in the recipient cell.
Figure 9.27 Transfer of chromosomal DNA by conjugation.

Transfer of the integrated F plasmid from an Hfr strain results in the cotransfer of chromosomal DNA because it is linked to the plasmid. The steps in transfer are similar to those shown in Figure 9.24a. However, the recipient remains F− and receives a linear fragment of donor chromosome attached to part of the F plasmid. For donor DNA to survive, it must be recombined into the recipient chromosome after transfer (not shown).
Because several distinct insertion sequences are present on the E. coli chromosome, a number of different Hfr strains are possible. A given Hfr strain always donates genes in the same order, beginning at the same position. However, Hfr strains that differ in the chromosomal integration site of the F plasmid transfer their genes in different orders (Figure 9.28). At some insertion sites, the F plasmid is integrated with its origin pointing in one direction, whereas at other sites the origin points in the opposite direction. The orientation of the F plasmid determines which chromosomal genes enter the recipient cell first, illustrating how virtually any gene on the chromosome can be mobilized by one Hfr configuration or another (Figure 9.28).
Figure 9.28 Formation of different Hfr strains.

Different Hfr strains donate genes from different origins and thus genes are transferred in different orders. (a) F plasmids can be inserted into various insertion sequences on the bacterial chromosome, forming different Hfr strains. (b) Order of gene transfer for different Hfr strains. See Figures 9.25–9.27 for details of Hfr formation and DNA transfer.
Transfer of Chromosomal Genes to the F Plasmid
Occasionally, integrated F plasmids may be excised from the chromosome. During excision, chromosomal genes may sometimes be incorporated into the liberated F plasmid. This can happen because both the F plasmid and the chromosome contain multiple identical insertion sequences where recombination can occur (Figure 9.25). F plasmids containing chromosomal genes are called F′ (F-prime) plasmids. When F′ plasmids promote conjugation, they transfer the chromosomal genes they carry at high frequency to the recipients. F′-mediated transfer resembles specialized transduction (Section 9.7) in that only a restricted group of chromosomal genes is transferred by any given F′ plasmid. Transferring a known F′ into a recipient allows one to establish diploids (two copies of each gene) for a limited region of the chromosome. Such partial diploids (merodiploids) are useful for genetic complementation experiments (Section 9.5).
We segue now to the final part of this chapter in which we explore archaeal genetics, movable genetic elements, and a major mechanism prokaryotic cells have evolved to protect their genomic integrity, a mechanism that has been exploited to revolutionize biology.
Check Your Understanding
In conjugation involving the F plasmid of Escherichia coli, how is the host chromosome mobilized?
Why does an Hfr×F− mating not yield two Hfr cells?
At which sites in the chromosome can the F plasmid integrate?
III: Gene Transfer in Archaea and Other Genetic Events
III: Gene Transfer in Archaea and Other Genetic Events
III Gene Transfer in *Archaea* and Other Genetic Events
**Although phylogenetically distinct from Bacteria, Archaea have evolved similar mechanisms for horizontal gene transfer. Some genetic elements can move about the chromosome, and a host of surveillance systems are present in prokaryotic cells that function to “search and destroy” foreign DNA.**
Although studies of the genetics of Archaea lag behind genetics research in Bacteria, they are showing progress, along with the development of archaeal versions of the tools necessary for detailed genetic analyses. In addition, some other genetic events in Bacteria reveal important genetic concepts even though they do not involve horizontal gene flow per se. These events can trigger genomic duplications and rearrangements, which increase genetic diversity (Chapter 13).
9.10 Horizontal Gene Transfer in *Archaea*
Although Archaea contain a single circular chromosome (Figure 9.29), as do most Bacteria (Section 6.2), and genome analysis clearly shows that horizontal transfer of archaeal DNA occurs in nature, experimental gene transfer systems are not as well developed for Archaea as they are for Bacteria. Some reasons for this are of a practical nature including the fact that most well-studied Archaea are extremophiles, capable of growth only under conditions of high salt or high temperature (Chapter 17). The temperatures necessary to culture some hyperthermophiles, for example, will melt agar, and alternative materials are required to form solid media and obtain colonies.
Figure 9.29 An archaeal chromosome, as shown by transmission electron microscopy.

The circular chromosome is from the hyperthermophile Sulfolobus, a member of the Archaea.
Another problem is that most known antibiotics do not affect Archaea. For example, penicillins do not affect Archaea because their cell walls lack peptidoglycan. The choice of selectable markers for use in genetic crosses is therefore often limited. However, novobiocin (a DNA gyrase inhibitor) and mevinolin (an inhibitor of isoprenoid biosynthesis) have been used to inhibit growth of extreme halophiles, and puromycin and neomycin (both protein synthesis inhibitors) inhibit methanogens. Auxotrophic strains (Section 9.1) of a few Archaea have also been isolated for genetic selection purposes.
Examples of Archaeal Genetics
No single species of Archaea has become a model organism for archaeal genetics, although more genetic work has been done on select species of extreme halophiles (Halobacterium, Haloferax, Section 17.1) than on any other Archaea. Various mechanisms of gene transfer have been discovered in select species of Archaea, including transformation, transduction, and conjugation. In addition, several plasmids are known and some have been used to construct cloning vectors, allowing genetic analysis through gene cloning and sequencing rather than through traditional genetic means, such as genetic crosses (Section 9.5). Transposon mutagenesis (Section 9.11) has been well developed in certain methanogens, including species of Methanococcus and Methanosarcina, and other tools such as shuttle vectors and other in vitro methods of genetic analysis (Chapter 12) have been developed for study of the unique biochemistry of the methanogens (Sections 14.15 and 17.2).
Archaeal Competence and Viral Transfer
Transformation occurs in the methanogen Methanococcus voltae and the hyperthermophiles Thermococcus kodakarensis and Pyrococcus furiosus, all of which are naturally competent (Section 9.6). Intraspecies exchange of plasmids occurs in some species of Thermococcus and Halorubrum through the budding of the cell envelope, a process that results in DNA-containing membrane vesicles as vehicles of genetic exchange. Formation of membrane vesicles occurs in species of all three domains of life, and these structures can carry both protein and nucleic acid. While DNA transfer by membrane vesicles has also been observed in Bacteria, the mechanism observed in transferring the pR1SE plasmid by the extreme halophile Halorubrum lacusprofundi blurs the line between transformation and transduction. While pR1SE does not encode genes of viral origin, the plasmid does contain genes encoding membrane proteins that are inserted into the host’s membrane; these are then used to form regularly shaped membrane vesicles that function to transfer the plasmid between cells (Figure 9.30). Thus, pR1SE mimics a virus because the genetic element encodes a portion of a structurally defined particle that transfers the plasmid between H. lacusprofundi strains.
Figure 9.30 Membrane vesicles and plasmid transfer in *Halorubrum lacusprofundi* R1S1.

(a) Transmission electron micrograph showing an isolated cell with membrane vesicles budding from the cell surface. Filaments around the cell are cell appendages and archaella. (b) Membrane vesicles containing plasmid DNA isolated from the supernatant of R1S1 cells. Each vesicle is 80–110 nm in diameter. Images modified from Erdmann, S., Tschitschko, B., Zhong, L., Raftery, M.J., and Cavicchioli, R. 2017. Nat. Microbiol. 2 : 1446.
Other conditions for transformation work reasonably well in several Archaea although the details vary from organism to organism. One approach requires removal of divalent metal ions, which in turn results in the partial disassembly of the glycoprotein cell wall layer that surrounds many archaeal cells (the S-layer, Section 2.5), and this allows access to transforming DNA. However, Archaea with rigid cell walls have proven difficult to transform, although electroporation (Section 9.6) sometimes works. One exception is in Methanosarcina species, organisms with a thick polysaccharide cell wall (Section 17.2). In this organism, a high-efficiency transformation system has been developed that employs DNA-loaded lipid preparations (liposomes) that are able to traverse the cell wall and deliver DNA into the cell.
Although viruses that infect Archaea are plentiful, transduction is rare. Only one archaeal virus, which infects the thermophilic methanogen Methanothermobacter thermautotrophicus, has been shown to transduce host genes. Unfortunately, the low burst size (about six phage virions liberated per cell) makes it impractical to use this system for gene transfer. Gene transfer agents (Section 9.7) have been found in one species of methanogen but do not appear to be widespread in Archaea.
Transfer by Cell-to-Cell Contact
Different types of conjugation have been detected in Archaea. Some strains of the thermophilic and acidophilic Sulfolobus solfataricus (Section 17.10) contain plasmids that promote conjugation between two cells in a manner similar to that seen in Bacteria. In this process, cell pairing is independent of pili formation, but as for Bacteria, DNA transfer is unidirectional. However, most of the genes encoding conjugation in S. solfataricus have little similarity to those in gram-negative Bacteria. An exception is a gene similar to the gene traG located on the Escherichia coli F plasmid (Sections 9.8 and 9.9); the protein product of the archaeal version of this gene participates in stabilizing mating pairs. Thus it is likely that the overall mechanism of conjugation in Archaea differs in significant ways from that in Bacteria.
Most species of Sulfolobus can also exchange DNA without the participation of a fertility plasmid. Such exchange is dependent on cell aggregation, a process that requires pili whose synthesis is triggered by UV radiation. While the exact mechanism of this unusual DNA exchange is unknown, these pili bring cells of the same species together through recognition of S-layer glycosylation patterns present on the cell walls.
Other Archaea form specialized structures between cells that allow for genetic exchange, and the formation of these structures is also independent of a fertility plasmid. For example, Figure 9.31 illustrates the formation of DNA-transferring nanotubes between cells of a Thermococcus species. Interestingly, unlike the other means of horizontal gene transfer discussed in this chapter, the Thermococcus nanotubes allow for bidirectional transfer of DNA. Thus, with both cells in an exchange able to function as recipients, the Thermococcus nanotubes likely facilitate very dynamic gene flow. Similarly, some halobacteria also form cytoplasmic bridges between mating cells that are used for DNA transfer. Although the nanotube and cytoplasmic bridge systems are not yet routine laboratory tools, they may well prove useful for the future development of more facile archaeal genetic transfer systems.
Figure 9.31 Nanotubes and *Thermococcus*.

Scanning electron micrograph showing nanotubes linking cells of a species of Thermococcus. A single Thermococcus cell is about 1 μm in diameter.
Check Your Understanding
Why is it usually more difficult to select recombinants with Archaea than with Bacteria?
9.11 Mobile DNA: Transposable Elements
As we have seen, molecules of DNA may move from one cell to another, but to a geneticist, the phrase “mobile DNA” has a special meaning. Mobile DNA refers to discrete segments of DNA that move as units from one location to another within other DNA molecules.
Although the DNA of certain viruses can be inserted into and excised from the genome of the host cell (Section 5.6 and Section 11.4), most mobile DNA consists of transposable elements. These are stretches of DNA that can move from one site to another. However, transposable elements are always found inserted into another DNA molecule such as a plasmid, a chromosome, or a viral genome. Transposable elements do not possess their own origin of replication. Instead, they are replicated when the host DNA molecule into which they are inserted is replicated.
Mastering Microbiology
Activities: Microbiology Animation: Transposons: Insertion Sequences
Transposable elements move by transposition, a process that is important in both genome rearrangement and genetic analysis. Transposable elements are extremely abundant and widespread in nature and are present in the genomes of all three domains of life and in many viruses and plasmids, suggesting that the elements offer a selective advantage by accelerating genome rearrangement and gene duplication (Section 13.8).
The two major types of transposable elements in Bacteria are insertion sequences (ISs) and transposons. Both elements have two important features in common: They carry genes encoding transposase, the enzyme necessary for transposition, and they have short inverted terminal repeats at their ends that are also needed for transposition (the ends of transposable elements are not free but are continuous with the host DNA molecule into which the transposable element has inserted). Figure 9.32 shows genetic maps of two well-studied transposable elements: the insertion element IS2 and the transposon Tn5.
Figure 9.32 Maps of the transposable elements IS*2* and Tn*5*.

The arrows above the maps show the direction of transcription of any genes on the elements. The gene encoding the transposase is tnp. (a) IS2 is an insertion sequence of 1327 bp with inverted repeats of 41 bp at its ends. (b) Tn5 is a composite transposon of 5.7 kbp containing the insertion sequences IS50L and IS50R at its left and right ends, respectively. IS50L is not capable of independent transposition because there is a nonsense mutation, marked by a blue cross, in its transposase gene. The genes kan, str, and bleo confer resistance to the antibiotics kanamycin (and neomycin), streptomycin, and bleomycin.
Insertion Sequences and Transposons
Insertion sequences (ISs) are the simplest type of transposable element. They are short DNA segments, about 1000 nucleotides long, and typically contain inverted repeats of 10–50 base pairs. Each different IS has a specific number of base pairs in its terminal repeats, and the only protein encoded is the transposase. Several hundred distinct IS elements have been characterized. IS elements are found in the chromosomes and plasmids of both Bacteria and Archaea, as well as in certain bacteriophages. Individual strains of the same bacterial species vary in the number and location of the IS elements they harbor. For instance, the genome of one strain of Escherichia coli has five copies of IS2 and five copies of IS3. Many plasmids, such as the F plasmid, also carry IS elements. Indeed, integration of the F plasmid into the E. coli chromosome is facilitated by recombination between identical IS elements on the F plasmid and the chromosome (Section 9.9 and Figure 9.25).
Transposons are larger than IS elements but have the same two essential components: inverted repeats at both ends and a gene encoding the transposase (Figure 9.32b). The transposase recognizes the inverted repeats and moves the segment of DNA flanked by them from one site to another. Consequently, any DNA that lies between the two inverted repeats is moved and is, in effect, part of the transposon. Genes included inside transposons vary widely. Some of these genes, such as antibiotic resistance genes, confer important new properties on the organism. Because antibiotic resistance is both important and easy to detect, most well-studied transposons contain antibiotic resistance genes as selectable markers. Examples include transposon Tn5, which encodes resistance to the antibiotics kanamycin (and neomycin), streptomycin, and bleomycin (Figure 9.32b), and Tn10, which encodes tetracycline resistance.
Because any genes lying between the inverted repeats become part of a transposon, it is possible to get hybrid transposons that display complex behavior. For example, conjugative transposons contain tra genes (Figures 9.25 and 9.26) and can therefore move between bacterial species by conjugation as well as transpose from place to place within a bacterial genome.
Mechanisms of Transposition
Both the inverted repeats (located at the ends of transposable elements) and transposase are essential for transposition. The transposase recognizes, cuts, and ligates the DNA during transposition. When a transposable element is inserted into target DNA, a short sequence in the target DNA at the site of integration is duplicated during the insertion process (**Figure 9.33*a***). The duplication arises because single-stranded DNA breaks are made by the transposase. The transposable element is then attached to the single-stranded ends that have been generated. Finally, enzymes of the host cell repair the single-strand portions, which results in the duplication.
Figure 9.33 Transposition.
(a) Insertion of a transposable element generates a duplication of the target sequence. Note the presence of inverted repeats (IR) at the ends of the transposable element. (b) Two mechanisms of transposition. Donor DNA (carrying the transposon) is shown in green, and recipient DNA carrying the target sequence is shown in yellow. In both conservative and replicative transposition, the transposase inserts the transposon (purple) into the target site (red) on the recipient DNA. During this process, the target site is duplicated. In conservative transposition, the donor DNA is left with a double-stranded break at the previous location of the transposon. In contrast, after replicative transposition, both donor and recipient DNA possess a copy of the transposon.
Two mechanisms of transposition are known: conservative and replicative (Figure 9.33b). In conservative transposition, as occurs with the transposon Tn5 (Figure 9.32b), the transposon is excised from one location and reinserted at a second location. The copy number of a conservative transposon therefore remains at one. By contrast, during replicative transposition, a new copy of the transposon is produced and inserted at the second location. Thus, after a replicative transposition event, one copy of the transposon remains at the original site, while a second copy is incorporated at the new site.
Utility of Transposon Mutagenesis
When a transposon inserts itself within a gene, the DNA sequence in the gene is altered and a mutation occurs (Figure 9.34). Mutations due to transposon insertion do occur naturally. However, laboratory use of transposons has been a powerful genetic tool to create a library of bacterial mutants. To do this, transposons carrying antibiotic resistance genes are used. With conjugation or transformation as the transfer mechanism, the transposon is introduced into the target cell on a plasmid that cannot replicate in that particular host. Consequently, antibiotic-resistant colonies will mostly be due to insertion of the transposon into the bacterial genome.
Figure 9.34 Transposon mutagenesis.

The transposon moves into the middle of gene 2. Gene 2 is now disrupted by the transposon and is inactivated. Gene A from the transposon is now expressed from the chromosome.
Because bacterial genomes contain relatively little noncoding DNA, most transposon insertions will occur in genes that encode proteins. This technique can be used to determine the function of a novel gene if a screening method is available. For example, if a transposon inserts into a gene encoding a product required for biofilm formation (biofilms are colonies of microbes encased in an adhesive and attached to a surface, Section 4.9), the transposon mutant will no longer grow in the biofilm mode. Then, further genetic analyses can be performed to reveal which gene the transposon has disrupted. Figure 9.35 illustrates the use of transposon mutagenesis to study flagella structure and function in a notorious biofilm-producing bacterium, Pseudomonas aeruginosa (Section 8.10). In this research, transposon mutagenesis combined with a special staining technique was used to identify the protein FlgK of the flagellar hook (Section 2.9) as a protein required for biofilm formation.
Figure 9.35 Utility of transposon mutagenesis.

A transposon (Tn) conferring kanamycin resistance (KmR) is added to a culture of wild-type Pseudomonas aeruginosa. After selection for integration of the transposon using the antibiotic kanamycin, the mutants are screened for biofilm formation in a microtiter plate. Cultures that produce biofilm adhere to the microtiter plate (ring in left tube of inset photo) and the resulting biofilm can be stained with crystal violet. Primers specific to the Tn can be used to determine the Tn insertion site and the identity of the interrupted gene. flgK encodes a protein in the flagellar hook.
Two transposons widely used for mutagenesis of E. coli and related bacteria are Tn5 (Figure 9.32b) and Tn10. Many Bacteria, a few Archaea, and the yeast Saccharomyces cerevisiae have all been mutagenized using genetically engineered transposons. In Chapter 10 we will learn about how engineered transposons can be used in metagenomic studies to determine the role of individual genes in a microbe’s overall fitness. More recently, transposons have even been used to isolate mutations in animals, including mice.
We finish this chapter with a consideration of genomic stability (in contrast to the genomic alterations we have discussed thus far) and preview the application of this process to biotechnology and synthetic biology.
Check Your Understanding
Which features do insertion sequences and transposons have in common?
What is the significance of the terminal inverted repeats of transposons?
How can transposons be used in bacterial genetics?
9.12 Preserving Genomic Integrity and CRISPR
While DNA transfer is key to the diversity and evolution of Bacteria and Archaea, cells also have mechanisms to prevent horizontal gene transfer and viral infection from occurring. With viruses outnumbering prokaryotic cells in nature by a factor of 10, barriers against gene transfer are key to preserving genome integrity and ultimately species maintenance. Figure 9.36 provides an overview of gene transfer barriers that work in concert in both Bacteria and Archaea. While our focus here is on mechanisms to prevent DNA transfer from viruses, such as modification of viral host cell receptors (Section 5.5 and Figure 9.36), these barriers also target incoming naked and plasmid DNA.
Figure 9.36 Barriers to horizontal gene transfer in *Bacteria* and *Archaea* using phage infection as an example.

Some possible outcomes of viral attack: (a) Successful viral infection. (b) The virus is prevented from recognizing and binding to its host cell; viral DNA cannot enter. (c) The foreign DNA is degraded by host cell restriction endonucleases (host cell DNA is not degraded because it is chemically modified at restriction cut sites). (d) The foreign DNA is modified by the host cell’s replication machinery such that it is not replicated. (e) Programmed cell death kills the host and thus prevents successful infection. (f) CRISPR activity destroys foreign DNA: crRNAs and Cas proteins detect DNA as previous invading DNA and degrade it through nuclease activity. Abbreviations: RE, restriction endonuclease; ME, modification enzyme; crRNA, CRISPR (clustered regularly interspaced short palindromic repeat) RNA; Cas (CRISPR-associated) protein.
Because barriers to horizontal gene transfer must distinguish invading DNA from self DNA, and protect the latter, they are considered bacterial and archaeal immune systems. Thus, processes that are nonspecific are considered analogous to innate immunity (Chapter 26), whereas mechanisms that target specific incoming nucleic acid are considered a form of adaptive immunity (Chapter 27). Despite engaging these forms of bacterial and archaeal immunity, the relationship between viruses and their bacterial and archaeal hosts is not static but instead extremely dynamic. That is, although microbial cells possess several weapons to battle viral attack, viruses counter these immune systems with weapons of their own, triggering a microbial “arms race” for cell survival and viral propagation.
Bacterial and Archaeal Innate Immunity
In our discussion of viruses in Section 5.5, we briefly considered restriction endonucleases—proteins produced by both Bacteria and Archaea that target and destroy foreign DNA—and how these enzymes protect the cell from viral attack. Restriction enzymes also target foreign DNA entering the cell as a result of transformation and conjugation events (Figure 9.36). The cell’s DNA is protected from its own restriction endonucleases because the enzymes’ recognition sequences are chemically modified (Section 12.2). As we mentioned in Chapter 5, some viruses avoid restriction endonuclease surveillance by modifying their DNA to mimic the chemical modification found in the host genome. In response, some bacteria have evolved altered restriction enzyme systems that recognize this viral DNA modification and still degrade the incoming viral DNA, preventing viral infection.
An example of this microbial arms race is that of T4 bacteriophage and its host cell Escherichia coli. The adaptation of E. coli recognizing modified phage DNA has now selected for T4 bacteriophages that modify their DNA in a different manner—by glycosylating (adding sugars) to specific DNA bases. Then, as one might expect in an expanding arms race, some strains of E. coli have evolved restriction systems that recognize glycosylated viral DNA and destroy it. As a countermeasure to this, some T4 strains have evolved a protein that inhibits these modified restriction enzymes. In a constant attempt to stay ahead, other E. coli strains have evolved endonucleases that are not inhibited by these bacteriophage proteins, and so it goes, back and forth as both predator and prey fight to survive and propagate in the face of “the enemy.”
Recent studies have uncovered yet additional mechanisms for thwarting viral infection, such as the phage exclusion and abortive infection systems present in both Bacteria and Archaea (Figure 9.36). Phage exclusion systems are a variant of restriction enzyme systems. Genes of the phage exclusion system encode enzymes that recognize and then modify incoming foreign DNA so that it cannot be replicated; this modification results in replication of only the host cell’s DNA and not the modified DNA. In contrast, abortive infection systems trigger the host cell to commit suicide (Figure 9.36). These systems are encoded by toxin–antitoxin systems (Section 8.12) that result in programmed cell death. Although the host cell dies during this abortive infection, its death prevents more virions from being produced and released to infect other cells in the population, thereby increasing the survival rate in the host population.
Adaptive Immunity and CRISPR
A major antiviral defense system present in both Bacteria and Archaea is CRISPR—clustered regularly interspaced short palindromic repeats—which functions to seek out and destroy foreign nucleic acid. While some CRISPR systems can target RNA viruses, our discussion will focus on systems that target foreign DNA. Besides protecting against viral infection, CRISPR also helps the cell maintain the stability and integrity of its genome by destroying certain plasmids and other genes obtained from horizontal transfers.
CRISPR regions are short repeats of constant DNA sequence present on the host chromosome that alternate with short variable DNA sequences called spacers (Figure 9.37). Spacers correspond to sequences of viral or other foreign DNA and function as a “memory bank” of past encounters with a virus in a manner reminiscent of how animals produce antibodies and long-lasting memory cells against an infecting virus. Because of this specificity, the CRISPR system is analogous to the adaptive immune response of vertebrate animals (Chapter 27).
Figure 9.37 CRISPR defense system against viruses and foreign DNA.

(a) Immunization. Incoming viral DNA is targeted by the memorizing complex of Cas proteins. This complex selects a protospacer region based on protospacer adjacent motif (PAM) sequences located on the viral genome. Once the protospacer is excised from the viral genome, the memorizing complex inserts the protospacer into the CRISPR region of the chromosome, resulting in a unique spacer region. (b) Interference. The chromosomal CRISPR region is transcribed and processed by Cas proteins into crRNAs that correspond to the individual spacer regions. Each spacer segment corresponds to previous encounters with incoming foreign nucleic acid. Cas proteins bind to these crRNAs and search for complementary DNA. If a crRNA binds to the DNA of an invading virus, forming a crRNA:DNA duplex, the endonuclease activity of the cleavage complex is triggered and results in the degradation of incoming viral DNA.
Besides the spacer regions, another essential component of the CRISPR system is the Cas (CRISPR-associated) proteins. These proteins possess endonuclease activity and both mediate the defense against foreign DNA and incorporate new spacers into the CRISPR region. When a virus attaches to a host cell and injects its DNA, the Cas proteins of a CRISPR region may recognize specific DNA sequences known as protospacer adjacent motifs (PAMs) (Figure 9.37a). The Cas protein then cleaves the viral DNA at a locus near this PAM (termed the protospacer) and inserts the short DNA region into the CRISPR region of the chromosome, where it becomes a spacer (Figure 9.37a). The insertion of a spacer into the CRISPR region confers “genetic memory” (referred to as immunization) on the cell and sets the stage for later encounters with the same virus.
When an immunized cell encounters the same virus later, the Cas proteins quickly destroy the incoming DNA in an RNA-dependent process. While the genomic CRISPR region does not contain open reading frames, it does have a promoter (Section 6.5). The resulting transcript is considered the pre-CRISPR RNA (pre-crRNA) and contains an array of RNA sequences complementary to both the repeat and spacer regions. The Cas proteins then process the transcript into individual spacer RNAs by targeting the repeat regions (Figure 9.37b). These crRNAs then associate with Cas proteins within the cleavage complex and begin surveillance for complementary incoming viral DNAs. Any viral DNA:crRNA duplexes formed are cleaved by the endonuclease activity of Cas proteins, and the invading DNA is degraded in a process called interference (Figure 9.37b). With part of its DNA destroyed in this way, the foreign DNA molecule of an invading virus cannot proceed to replicate and the infection (and threat to the cell) is thwarted.
One of the major conundrums of the CRISPR system is how a host can survive the initial viral invasion long enough to become immunized. For the CRISPR system to successfully prevent viral infection, a spacer corresponding to a region of the viral genome must already be present in the CRISPR locus. Immunization probably occurs when an incoming virus has been inactivated by environmental factors (such as ultraviolet radiation) or when the host’s restriction enzyme system cleaves the invading DNA before a successful infection can be initiated.
Distribution of CRISPR Systems and Anti-CRISPR Systems
CRISPR systems are widely distributed in both Archaea and Bacteria. Approximately 90% of the sequenced genomes of Archaea and 70% of those of Bacteria possess a CRISPR system. The utility of the system was first demonstrated in the dairy industry where starter cultures used for milk fermentation were susceptible to rampant bacteriophage infection. However, a strain of Streptococcus thermophilus was found to be resistant to virulent bacteriophage, and the difference between this S. thermophilus strain and those susceptible to viral infection was the spacers within its CRISPR region.
Because of the dynamic nature of the phage–host interaction, viruses have evolved mechanisms to avoid CRISPR surveillance and destruction. These include mutation of the PAM regions recognized by the memorizing complex of Cas proteins and the production of proteins that inhibit activity of the cleavage complex of Cas proteins (Figure 9.37b). This illustrates how the CRISPR system—central as it may be to preventing attacks on a cell’s genome—does have a drawback in its reliance on recognizing previously encountered DNA sequences that can continue to mutate in their host and then reappear in the future in a form not recognized by CRISPR.
In addition, a phage-encoded (in contrast to cell-encoded) CRISPR system has been identified in the genome of a bacteriophage that infects Vibrio cholerae, the bacterium that causes cholera. The phage-encoded crRNAs target genes in V. cholerae that encode a defense system that prevents bacteriophage propagation; when these genes are inactivated, the cell’s defense system is inoperative, a rather elaborate example of the “arms race” between bacteria and their viruses.
While CRISPRs are a major weapon Bacteria and Archaea have evolved for viral surveillance and maintaining genomic integrity, the mechanisms employed by CRISPR/Cas have not been overlooked by the scientific community. In Chapter 12 we discuss how the CRISPR/Cas system has been exploited as a powerful tool for synthetic biology and has opened up the possibility of radically new therapeutic strategies for human medicine (Section 12.13).
Check Your Understanding
Describe two ways that bacterial and archaeal cells can avoid the entry of foreign DNA. How can viruses overcome these defenses?
Why is the CRISPR system considered a prokaryotic “adaptive immune system”?
How do the crRNAs and Cas proteins protect the cell from invading DNA?
Chapter Review
Go to Mastering Microbiology for videos, animations, practice tests, and more.
I Mutation
9.1 Mutation is a heritable change in the nucleotide sequence of the genome and may lead to a change in phenotype. Selectable mutations are those that give the mutant a growth advantage under certain environmental conditions and are especially useful in genetic research. If selection is not possible, mutants must be identified by screening.
Q Write a one-sentence definition of the term “genotype.” Do the same for “phenotype.” Does the phenotype of an organism automatically change when a change in genotype occurs? Why or why not? Can phenotype change without a change in genotype? In both cases, give examples to support your answer.
9.2 Mutations, either spontaneous or induced, are in the base sequence of the nucleic acid in a genome. A point mutation is due to a single base-pair change. In a nonsense mutation, the codon becomes a stop codon and an incomplete polypeptide is made. Deletions and insertions cause more dramatic changes in the DNA, including frameshift mutations that often result in complete loss of gene function.
Q What are silent mutations? From your knowledge of the genetic code, why do you think most silent mutations affect the third position in a codon?
9.3 Different types of mutations occur at different frequencies. For a typical bacterium, mutation rates of 10−6 to 10−7 per kilobase pair are generally seen. Although RNA and DNA polymerases introduce sequence errors at about the same rate, RNA genomes typically accumulate mutations at much higher frequencies than DNA genomes.
Q What is the difference between same-site and second-site revertants?
9.4 Mutagens are chemical, physical, or biological agents that increase the mutation rate. Mutagens can alter DNA in many different ways. However, alterations in DNA are not mutations unless they are inherited. Some DNA damage can lead to cell death if not repaired, and both error-prone and high-fidelity DNA repair systems exist.
Q Give an example of one biological, one chemical, and one physical mutagen and describe the mechanism by which each causes a mutation.
II Gene Transfer in *Bacteria*
9.5 Homologous recombination occurs when closely related DNA sequences from two distinct genetic elements are combined together in a single element. Recombination is an important evolutionary process, and cells have specific mechanisms for ensuring that recombination takes place.
Q What are heteroduplex regions of DNA, and what process leads to their formation?
9.6 Certain bacteria exhibit competence, a state in which cells are able to take up free DNA released by other bacteria, a process called transformation. Once this DNA has been taken up, typically through the action of pili, incorporation of donor DNA into a recipient cell requires the activity of the single-strand binding protein RecA and several other enzymes. Only competent cells are transformable.
Q Explain why recipient cells do not successfully take up plasmids during natural transformation.
9.7 Transduction is the transfer of host genes from one bacterium to another by a bacterial virus. In generalized transduction, defective virus particles randomly incorporate fragments of the cell’s chromosomal DNA, but the transducing efficiency is low. In specialized transduction, the DNA of a temperate virus excises incorrectly and takes adjacent host genes along with it; the transducing efficiency here may be very high.
Q Explain how a generalized transducing particle differs from a specialized transducing particle.
9.8 Conjugation is a mechanism of DNA transfer in Bacteria and Archaea that requires cell-to-cell contact. Conjugation is controlled by genes carried by certain plasmids (such as the F plasmid) and requires transfer of the plasmid from a donor cell to a recipient cell. Plasmid DNA transfer requires replication using the rolling circle mechanism.
Q What is a conjugative pilus and which cell type, F− or F+, would produce this structure?
9.9 The donor cell chromosome can be mobilized for transfer to a recipient cell. This requires an F plasmid to integrate into the chromosome to form the Hfr phenotype. Because transfer of the integrated F plasmid is rarely complete, recipient cells rarely become F+. F′ plasmids are previously integrated F plasmids that have excised and captured some chromosomal genes.
Q What is a merodiploid, and how does an F′ plasmid yield a merodiploid?
III Gene Transfer in *Archaea* and Other Genetic Events
9.10 Archaeal research lags behind bacterial research in the development of systems for gene transfer. Many antibiotics are ineffective against Archaea, making it difficult to select recombinants effectively, and the unusual growth conditions needed by many Archaea also make genetic experimentation challenging. Nevertheless, the genetic transfer systems of Bacteria—transformation, transduction, and conjugation—are all known in Archaea.
**Q Explain one type of conjugation in Archaea and how it differs from F-plasmid-mediated conjugation.**
9.11 Transposons and insertion sequences are genetic elements that can move from one location on a host DNA molecule to another by transposition. Transposition can be either replicative or conservative. Transposons often carry genes encoding antibiotic resistance and can be used to identify the functions of unknown genes.
Q What are the major differences between insertion sequences and transposons?
9.12 Bacterial and archaeal cells employ various mechanisms to prevent gene transfer and viral infection. However, the fast mutation rate of viruses allows them to adapt to these strategies. The clustered regularly interspaced short palindromic repeat (CRISPR) system is an RNA-based immune system that protects the genome of Bacteria and Archaea from invading DNA resulting from viral infection or conjugation. If small RNA molecules resulting from the spacer regions of the CRISPR region bind to incoming complementary DNA, Cas proteins destroy the nucleic acid duplex.
Q Explain why incoming DNA recognized by a short RNA molecule expressed from the CRISPR region cannot be completely foreign to the cell.
Application Questions
A constitutive mutant is a strain that continuously makes a protein that is inducible in the wild type. Describe two ways in which a change in a DNA molecule could lead to the emergence of a constitutive mutant. How could these two types of constitutive mutants be distinguished genetically?
Although a large number of mutagenic chemicals are known, none is known that induces mutations in only a single gene (gene-specific mutagenesis). From what you know about mutagens, explain why it is unlikely that a gene-specific chemical mutagen will be found. How then is site-specific mutagenesis accomplished?
Why is it difficult in a single experiment to transfer a large number of genes to a recipient cell using transformation or transduction?
Transposable elements cause mutations when inserted within a gene. These elements disrupt the continuity of a gene. Introns also disrupt the continuity of a gene, yet the gene is still functional. Explain why the presence of an intron in a gene does not inactivate that gene but insertion of a transposable element does.
Chapter Glossary
an organism that has developed an additional nutritional requirement compared with the wild type, often as a result of mutation Conjugation
the transfer of genes from one prokaryotic cell to another by a mechanism requiring cell-to-cell contact Frameshift mutation
a mutation in which insertion or deletion of nucleotides changes the groups of three bases in which the genetic code is read within an mRNA, usually resulting in a faulty product Genotype
the complete genetic makeup of an organism; the complete description of a cell’s genetic information Heteroduplex
a DNA double helix composed of single strands from two different DNA molecules Hfr cell
a cell with the F plasmid integrated into the chromosome Horizontal gene transfer
the unidirectional transfer of genes between cells through a process uncoupled from reproduction Induced mutation
a mutation caused by external agents such as mutagenic chemicals or radiation Insertion sequence (IS)
the simplest type of transposable element, which carries only genes that participate in transposition Missense mutation
a mutation in which a single codon is altered so that one amino acid in a protein is replaced with a different amino acid Mutagen
an organism whose genome carries a mutation Mutation
a heritable change in the base sequence of the genome of an organism Nonsense mutation
a mutation in which the codon for an amino acid is changed to a stop codon Phenotype
the observable characteristics of an organism Point mutation
a mutation that involves a single base pair Recombination
a resorting or rearrangement of DNA fragments resulting in a new sequence Reversion
an alteration in DNA that reverses the effects of a prior mutation Rolling circle replication
a mechanism, used by some plasmids and viruses, of replicating circular DNA, which starts by nicking and unrolling one strand. For a single-stranded genome, this is preceded by using the still-circular strand as a template for DNA synthesis; for a double-stranded genome, the unrolled strand is used as a template for DNA synthesis Screening
a procedure that permits the identification of organisms by phenotype or genotype, but does not inhibit or enhance the growth of particular phenotypes or genotypes Selection
placing organisms under conditions that favor or inhibit the growth of those with a particular phenotype or genotype Silent mutation
a change in DNA sequence that has no effect on the phenotype SOS repair system
a DNA repair system activated by DNA damage Spontaneous mutation
a mutation that occurs “naturally” without the help of mutagenic chemicals or radiation Transduction
the transfer of host cell genes from one cell to another by a virus Transformation
the transfer of bacterial genes through the uptake of free DNA from the environment Transition
a mutation in which a pyrimidine base is replaced by another pyrimidine or a purine is replaced by another purine Transposable element
a genetic element able to move (transpose) from one site to another on host DNA molecules Transposon
a type of transposable element that carries genes in addition to those required for transposition Transversion
a mutation in which a pyrimidine base is replaced by a purine or vice versa Wild-type strain
a bacterial strain isolated from nature or one used as a parent in a genetics investigation
Unit 3 Genomics, Synthetic Biology, and Evolution
Chapter 13 Microbial Evolution and Genome Dynamics