16 Phylogenetic Diversity of Bacteria
## Chapter 16 Phylogenetic Diversity of Bacteria
**II Firmicutes, Tenericutes, and *Actinobacteria***
**IV Chlamydiae, Planctomycetes, and *Verrucomicrobia***
Bacterial Diversity and Human Health
Animals, particularly those that eat plants, have gut microbes that convert food into nutrients for the host. These animals lack enzymes for digesting the lignin and cellulose of plant cell walls and have evolved specialized organs that house microbes to perform these tasks. In humans, this organ is the colon, also known as the large intestine.
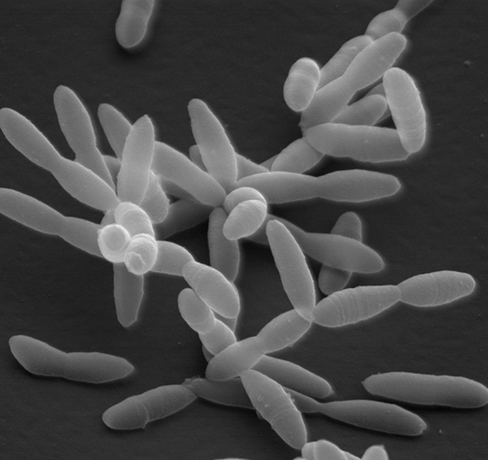
The average human colon contains hundreds of microbial species, and up to a hundred trillion microbial cells, many of which belong to the order Clostridiales within the phylum Firmicutes. The Clostridiales are a highly diverse group of gram-positive anaerobes that employ a fermentative metabolism. Clostridiales are crucial to gut function, and changes in their abundance and diversity can have important consequences for human health.
Christensenella minuta (see scanning electron micrograph), a member of the Clostridiales, was first discovered in human feces. C. minuta is a strictly anaerobic, nonsporulating and nonmotile fermentative bacterium that ferments sugars to short-chain fatty acids, H2, and CO2. C. minuta often co-occurs with methane-producing Archaea and may grow syntrophically with these H2-consuming organisms. Microbiome research suggests that C. minuta and other species of the family Christensenellaceae are indicators of a healthy gut. These bacteria are depleted in obese individuals as well as those suffering from metabolic syndrome, inflammatory bowel disease, Crohn’s disease, and ulcerative colitis. In addition, the introduction of live cells of C. minuta into mice reduces their body fat levels compared with control mice given heat-killed cells. Moreover, the relative abundance of Christensenellaceae in the gut is genetically determined in humans, with up to 40% of variation in their relative abundance attributable to host genetics.
These results show that our genes influence our microbial relationships and that our gut microbes can influence our health. A full understanding of our gut health therefore depends upon our understanding of gut bacterial diversity.
Source: Goodrich, J.K., et al. 2014. Human genetics shape the gut microbiome. Cell 159: 789.
In the previous chapters we examined the metabolic and ecological diversity of microorganisms. In this and the next two chapters, we shift our focus to the phylogenetic diversity of microorganisms. We considered the difference between metabolic, ecological, and phylogenetic diversity in Section 15.1. In this chapter, we examine the major lineages of Bacteria (**Figure 16.1*a***) and focus on the Archaea and microbial Eukarya in Chapters 17 and 18, respectively.
Figure 16.1 Some major phyla of *Bacteria* based on 16S ribosomal RNA gene sequence comparisons.

(a) Depicted are the major phyla of Bacteria that have cultivated species. The area of each wedge is roughly proportional to the number of described cultivated species in each group. Analyses of 16S rRNA gene sequences from natural environments suggest there are more than 80 bacterial phyla. (b) Numbers of cultured and characterized species (green bars) and known 16S rRNA gene sequences (phylotypes, red bars) for each of the 29 major bacterial phyla that have at least one type species in pure culture. Also shown are related data for the different classes of Proteobacteria. Differences between the size of the red and green bars indicate the degree to which members of each group are common in natural environments but difficult to cultivate in isolation. Note that the abscissa is a log scale.
Including phyla of Bacteria known only from 16S ribosomal RNA (rRNA) gene sequences retrieved from the environment (Sections 19.6 and 19.8), over 80 phyla can be distinguished. However, only about 30 of these contain species that have been characterized in laboratory culture (Figure 16.1b). Remarkably, more than 90% of cultivated genera and species of Bacteria originate in only four phyla: Proteobacteria, Actinobacteria, Firmicutes, and Bacteroidetes (Figure 16.1b).
With more than 12,000 species of bacteria described, we obviously cannot consider them all. Therefore, using phylogenetic trees to focus our discussion, we will explore some of the best-known species from a broad diversity of phyla. In this chapter, we will consider species from 20 bacterial phyla, focusing on those with the largest numbers of characterized species. We begin our tour of the Bacteria with the phylum Proteobacteria, a hotbed of cultured species in this domain.
I Proteobacteria
**The Proteobacteria are divided into six classes, the Alpha-, Beta-, Gamma-, Delta-, Epsilon-, and Zetaproteobacteria. Proteobacteria is the largest and most metabolically diverse phylum of Bacteria and leads in numbers of cultivated and well-characterized species.**
The Proteobacteria are by far the largest and most metabolically diverse phylum of Bacteria (Figure 16.2). More than a third of characterized species of Bacteria originate within this group (Figure 16.1b), and Proteobacteria constitute the majority of known bacteria of medical, industrial, and agricultural significance.
Figure 16.2 Phylogenetic tree and metabolic links of some key genera of *Proteobacteria*.

Phylogeny of representative genera of Proteobacteria as revealed by analysis of 16S rRNA gene sequences. Note how identical metabolisms are often distributed in phylogenetically distinct genera, suggesting that horizontal gene flow has been extensive in the Proteobacteria. Some organisms listed may have multiple properties; for example, some sulfur chemolithotrophs are also iron or hydrogen chemolithotrophs, and several of the organisms listed can fix nitrogen. Phylogenetic analyses were performed and the phylogenetic tree constructed by Marie Asao, Ohio State University.
As a group, the Proteobacteria are all gram-negative bacteria. They show an exceptionally wide diversity of energy-generating mechanisms, with chemolithotrophic, chemoorganotrophic, and phototrophic species (Figure 16.2). We have already encountered diverse representatives of this group in Chapters 14 and 15, where we considered the tremendous metabolic and ecological diversity of the Proteobacteria. Indeed, only a few types of metabolism do not occur within the Proteobacteria, including methanogenesis, which is only in Archaea (Section 17.2); oxygenic phototrophy, which is only in Cyanobacteria (Section 15.3); anammox, which is only in Planctomyces (Section 14.10); the complete oxidation of ammonia all the way to nitrate, a process called comammox (Section 21.3), which is only found in Nitrospirae (Section 16.21); and anaerobic methanotrophy (Section 14.16). The Proteobacteria are equally diverse in terms of their relationship to O2, with anaerobic, microaerophilic, and facultatively aerobic species known. Morphologically, they also exhibit a wide range of cell shapes, including straight and curved rods, cocci, spirilla, and filamentous, budding, and appendaged forms (Section 1.3 and Figure 1.8).
As we proceed through this chapter, we will need to recall the descending hierarchy of microbial systematics: domain (Bacteria or Archaea)→phylum→class→order→family→genus→species (Section 13.12 and Table 13.1). We will use these terms frequently in this chapter. Based on 16S rRNA gene sequences, the phylum Proteobacteria can be divided into six classes: Alpha-, Beta-, Gamma-, Delta-, Epsilon-, and Zetaproteobacteria. Each class contains many genera with the exception of the Zetaproteobacteria, which is composed of a single characterized species, the marine iron-oxidizing bacterium Mariprofundus ferrooxydans (Section 15.14).
Despite the phylogenetic breadth of the Proteobacteria, species in different classes often have similar metabolisms. For example, phototrophy and methylotrophy occur in three different classes of Proteobacteria, and nitrifying bacteria span four classes of Proteobacteria (Figure 15.1). This suggests that horizontal gene flow (Chapter 9 and Section 13.9) has played a major role in shaping the metabolic diversity of the Proteobacteria. The sharing of metabolic traits in the different classes of Proteobacteria is also a good reminder that phenotype and phylogeny often provide different views of prokaryotic diversity (Section 15.1).
16.1 Alphaproteobacteria
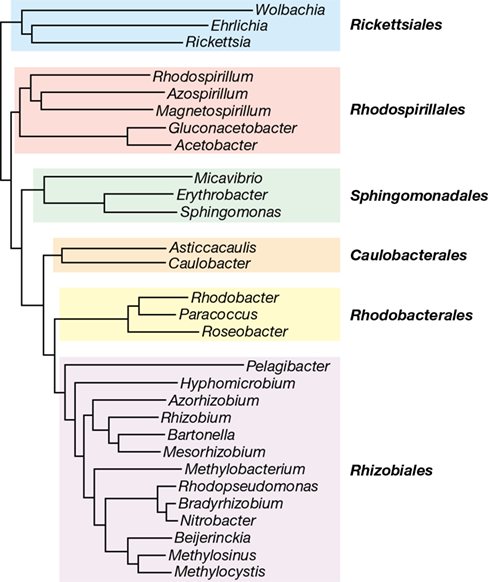
With about one thousand described species, the Alphaproteobacteria are the second largest class of Proteobacteria (Figure 16.1b). The Alphaproteobacteria contain extensive functional diversity (Figure 16.2, Figure 15.1), and many genera in this group have already been considered in Chapters 14 and 15. Most species are obligate aerobes or facultative aerobes and many are oligotrophic, preferring to grow in environments that have low nutrient concentrations. There are 10 well-characterized orders within the Alphaproteobacteria, but the vast majority of species fall within the Rhizobiales, Rickettsiales, Rhodobacterales, Rhodospirillales, Caulobacterales, and Sphingomonadales (Figure 16.3 and Table 16.1).
Figure 16.3 Major orders of *Proteobacteria* in the class *Alphaproteobacteria*.

The phylogenetic tree was constructed using 16S rRNA gene sequences from representative genera of Alphaproteobacteria. Order names are shown in bold.
Table 16.1 Notable genera of *Alphaproteobacteria*

Key Genera: *Bartonella, Methylobacterium, Pelagibacter, Rhizobium, Agrobacterium*
The Rhizobiales (Figure 16.3) are the largest and most metabolically diverse order of Alphaproteobacteria and contain phototrophs (e.g., Rhodopseudomonas), chemolithotrophs (e.g., Nitrobacter), symbionts (e.g., rhizobia), free-living nitrogen-fixing bacteria (e.g., Beijerinckia), a few pathogens of plants and animals, and diverse chemoorganotrophs. The group gets its name from the rhizobia, a polyphyletic collection of genera that form root nodules and fix nitrogen in symbiotic association with leguminous plants (Section 23.4).
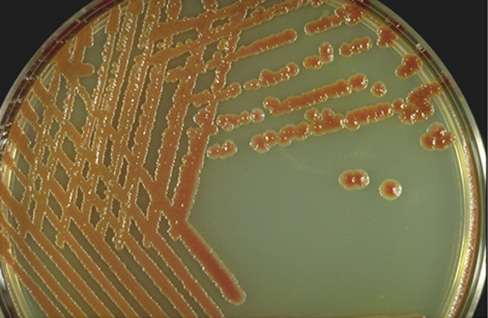
Among the Rhizobiales are nine genera that contain rhizobia: Bradyrhizobium, Ochrobactrum, Azorhizobium, Devosia, Methylobacterium, Mesorhizobium, Phyllobacterium, Sinorhizobium, and Rhizobium. These are typically chemoorganotrophs and obligate aerobes, and the genes that convey the ability to form root nodules have been distributed among these genera by horizontal gene transfer. Indeed, nodulation genes are found on large plasmids that can be transferred between cells (Section 23.4). Each rhizobial genus has a distinct range of plant hosts that can be colonized (Table 23.1). Rhizobia can be isolated by crushing nodules and spreading their contents on nutrient-rich solid media; colonies typically produce copious amounts of exopolysaccharide slime (Figure 16.4).
Figure 16.4 Colonies of *Rhizobium mongolense*.

Colonies of rhizobia often produce copious exopolysaccharide slime. These colonies of Rhizobium mongolense were grown on a medium low in nitrogen with sucrose as carbon source.
The organism Agrobacterium tumefaciens (also called Rhizobium radiobacter) is closely related to root nodule Rhizobium species but is a plant pathogen that causes crown gall disease (Section 23.6). A. tumefaciens is unable to form root nodules, and the genes that encode gall formation are also found on a plasmid. These pathogenicity genes, however, are unrelated to the plasmid-borne genes that mediate nodule formation in symbiotic rhizobia.
The genus Methylobacterium is one of the largest in the Rhizobiales. These species are often called “pink-pigmented facultative methylotrophs” because of the pink color of their colonies (Section 15.15) and their good growth on methanol. Species are commonly found on the surface of plants and in soils and freshwater systems. These organisms are also commonly encountered in toilets and baths where their growth on shower curtains, caulk, and in toilet bowls results in the formation of pink-pigmented biofilms. Species of Methylobacterium are readily isolated by pressing the surface of a plant leaf onto an agar Petri plate containing methanol as the sole source of carbon.
Bartonella is another notable genus of Rhizobiales. These organisms, once classified with the Rickettsiales, are intracellular pathogens of humans. Species of Bartonella can cause a variety of diseases in humans and other vertebrate animals. Bartonella quintana is the causative agent of trench fever, a disease that got its name because it decimated soldiers in the brutal trench warfare practiced in World War I. Other species of Bartonella can cause bartonellosis, cat scratch disease, and a variety of inflammatory diseases. These diseases are mediated by arthropod vectors including fleas, lice, and sand flies (Chapter 32). Species of Bartonella are fastidious and difficult to cultivate, and isolation is most commonly achieved using blood agar. When growing in tissue culture, cells of Bartonella grow on the outside surface of the eukaryotic host cells rather than within the cytoplasm or the nucleus.
Finally, the genus Pelagibacter also belongs to the Rhizobiales. Pelagibacter ubique is an oligotroph and an obligately aerobic chemoorganotroph that inhabits the photic zone of Earth’s oceans. This organism can make up 25% of the bacterial cells found at the ocean’s surface, and its numbers can reach 50% of cells in temperate waters in the summer; as a consequence, Pelagibacter ubique is likely the most abundant bacterial species on Earth (Section 20.12).
Figure 16.5 Rickettsias growing within host cells.

(a) Rickettsia rickettsii in tissue culture. Cells are about 0.3 μm in diameter. (b) Electron micrograph of cells of Rickettsiella popilliae within a blood cell of its host, the beetle Melolontha melolontha. The bacteria grow inside a vacuole within the host cell.
Species of the genus Rickettsia are the causative agents of several human diseases, including typhus (Rickettsia prowazekii) and spotted fever rickettsiosis, commonly called Rocky Mountain spotted fever (Rickettsia rickettsii)(Section 32.3). These organisms are closely associated with arthropod vectors and can be transmitted by ticks, fleas, lice, and mites. Most rickettsias are metabolically specialized, able to oxidize only the amino acids glutamate or glutamine and unable to oxidize glucose or organic acids. Rickettsias are unable to synthesize certain metabolites and must instead obtain them from host cells. Rickettsias do not survive long outside their hosts, and this may explain why they must be transmitted from animal to animal by arthropod vectors.
Electron micrographs of thin sections of rickettsial cells show a typical prokaryotic morphology including a cell wall (Figure 16.5b). The penetration of a host cell by a rickettsial cell is an active process, requiring both host and parasite to be viable and metabolically active. Once inside the host cell, the bacteria multiply primarily in the cytoplasm and continue replicating until the host cell is loaded with parasites (Figure 16.5; Figures 32.6 and 32.7). The host cell then bursts and liberates the bacterial cells.
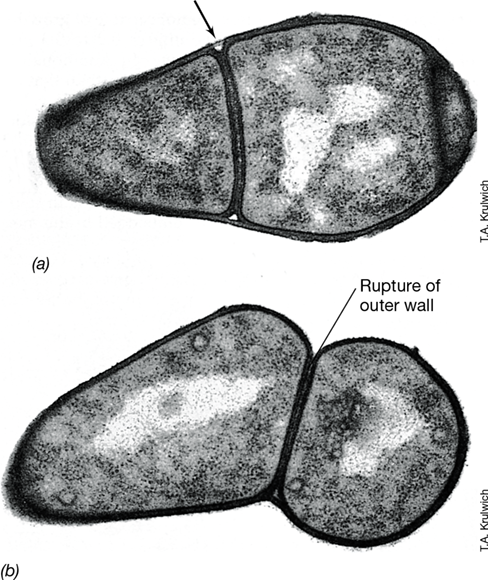
The genus Wolbachia contains intracellular parasites found within many arthropods and some nematodes (Figure 16.6). Species of Wolbachia infect an enormous diversity of insect species, and 10–70% of individual insects in a susceptible species carry Wolbachia. Wolbachia species can have any of several effects on their insect hosts. These include inducing parthenogenesis (development of unfertilized eggs), the killing of males, and feminization (the conversion of male insects into females). Wolbachia, because of their ability to alter host reproduction, have a major impact on the evolution of their insect hosts (Section 23.7).
Figure 16.6 *Wolbachia*.

Photomicrograph of a DAPI-stained egg of the parasitoid wasp Trichogramma kaykai infected with Wolbachia pipientis, which induces parthenogenesis. The W. pipientis cells are primarily located in the narrow end of the egg (arrows).
Wolbachia pipientis is the best-studied species in the genus. Cells of W. pipientis colonize the insect egg (Figure 16.6), where they multiply in vacuoles of host cells surrounded by a membrane of host origin. Wolbachia lack a cell wall and the ability to make peptidoglycan. Peptidoglycan can trigger host immune responses (Chapter 26), and the loss of genes for peptidoglycan synthesis helps this obligate intracellular organism escape host immune detection. Cells of W. pipientis are passed from an infected female to her offspring through infected eggs. Wolbachia-induced parthenogenesis occurs in a number of species of wasps. In the normal reproductive cycle of these insects, eggs that are unfertilized (and haploid) develop into males, while eggs that are fertilized (and diploid) develop into females. However, Wolbachia infection triggers a doubling of chromosome number in haploid eggs, and so an infected mother can only lay female eggs that contain her own DNA. Predictably, if female insects are fed antibiotics that kill Wolbachia, parthenogenesis ceases.
Key Genera: *Rhodobacter, Acetobacter, Caulobacter, Sphingomonas*
The orders Rhodobacterales and Rhodospirillales (Figure 16.3) contain metabolically diverse organisms that have been discussed previously, including purple nonsulfur bacteria (Rhodobacter and Rhodospirillum, Section 15.5), aerobic anoxygenic phototrophs (Roseobacter, Section 15.5), nitrogen-fixing bacteria (Azospirillum, Section 15.9), denitrifiers (Paracoccus, Section 15.10), and magnetotactic bacteria (Magnetospirillum, Section 15.20), among others.
The Caulobacterales are typically oligotrophic and strictly aerobic chemoorganotrophs. Species typically form prosthecae or stalks (Section 15.18), and many species display asymmetric forms of cell division. The characteristic genus is Caulobacter, which has a characteristic life cycle that we have discussed previously (Sections 8.8 and 15.18).
The Sphingomonadales include diverse aerobic and facultatively aerobic chemoorganotrophs as well as species of aerobic anoxygenic phototrophs (Erythrobacter) and a few obligate anaerobes. The characteristic genus is Sphingomonas, which consists of obligately aerobic and nutritionally versatile species. Sphingomonads are widespread in aquatic and terrestrial environments and are notable for their ability to metabolize a wide range of organic compounds including many aromatic compounds that are common environmental contaminants (toluene, nonylphenol, dibenzo-p-dioxin, naphthalene, and anthracene, among others). Consequently, sphingomonads have been widely studied as potential agents of bioremediation (Sections 22.4 and 22.5). These organisms are typically easy to cultivate and grow well on a variety of complex culture media.
Check Your Understanding
What are some ways in which Wolbachia species can affect insects? What evolutionary pressures have likely caused Wolbachia to lose the ability to make peptidoglycan?
Agrobacterium tumefaciens has been reclassified into the genus Rhizobium. What mechanism underlies the distribution of genes for virulence or symbiosis among the species of this genus?
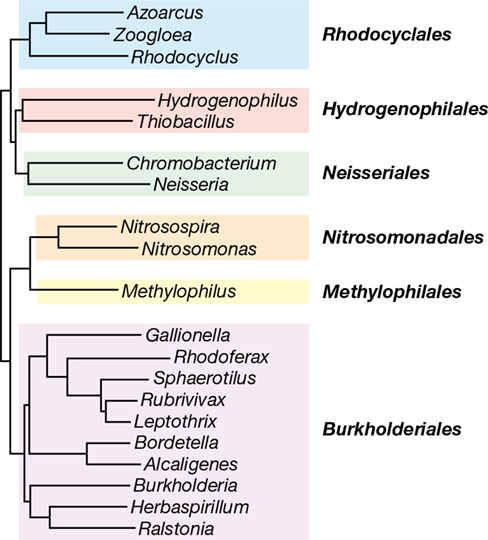
16.2 *Betaproteobacteria*
With about 500 described species, the Betaproteobacteria are the third largest class of Proteobacteria (Figure 16.1). The Betaproteobacteria contain an immense amount of functional diversity (Figure 16.2 and Figure 15.1), and many species in this group have already been considered in Chapter 15. A total of six orders of Betaproteobacteria have many characterized species: Burkholderiales, Hydrogenophilales, Methylophilales, Neisseriales, Nitrosomonadales, and Rhodocyclales (Figure 16.7), and we focus on these here.
Figure 16.7 Major orders of *Proteobacteria* in the class *Betaproteobacteria*.

The phylogenetic tree was constructed using 16S rRNA gene sequences from representative genera of Betaproteobacteria. Order names are shown in bold.
Key Genus: *Burkholderia*
The Burkholderiales contain species with a wide range of metabolic and ecological characteristics. Species include strictly aerobic, facultatively aerobic, and obligately anaerobic chemoorganotrophs, anoxygenic phototrophs, obligate and facultative chemolithotrophs, free-living nitrogen fixers, and pathogens of plants, animals, and humans.
Burkholderia is the type genus for the Burkholderiales. The genus Burkholderia includes diverse species of chemoorganotrophs with strictly respiratory metabolism. All species can grow aerobically, some also grow anaerobically with nitrate as the electron acceptor, and many strains are able to fix N2. The metabolic versatility of Burkholderia species with respect to organic compounds, and aromatic compounds in particular, has led to interest in their use in bioremediation (Section 22.4). Certain strains of Burkholderia have also been shown to promote plant growth. However, many species are potentially pathogenic for plants or animals. One of the best known of the pathogenic species is Burkholderia cepacia (Figure 16.8).
Figure 16.8 Colonies of *Burkholderia*.
Photograph of colonies of Burkholderia cepacia on an agar plate.
B. cepacia is primarily a soil bacterium but also an opportunistic pathogen. Often found in the rhizosphere of plants, B. cepacia can produce both antifungal and anti-nematodal compounds, and thus its ability to colonize plant roots can provide disease protection and promote plant growth. However, B. cepacia is also known as a plant pathogen in certain circumstances, and it is a major cause of soft rot in onions. B. cepacia has also emerged as an opportunistic hospital-acquired infection in humans, as it is a hardy organism that is difficult to eradicate from the clinical setting. B. cepacia can form secondary lung infections in patients who are immunocompromised or have pneumonia or cystic fibrosis. The ability of B. cepacia to form biofilms in the lung and its natural resistance to many antibiotics has made this organism, along with Pseudomonas aeruginosa, particularly dangerous for patients with cystic fibrosis (Section 8.10 and Section 20.4).
Key Genera: *Rhodocyclus, Zoogloea*
Like the Burkholderiales, the order Rhodocyclales contains species with diverse metabolic and ecological characteristics. The type genus for the Rhodocyclales is Rhodocyclus, a purple nonsulfur bacterium (Section 15.5). Like most purple nonsulfur bacteria, Rhodocyclus species grow best as photoheterotrophs but most can also grow as photoautotrophs with H2 as electron acceptor. Species can also grow by respiration in darkness, but they are typically found in illuminated anoxic environments where organic matter is present.
Zoogloea is another important genus of the Rhodocyclales. Zoogloea species are aerobic chemoorganotrophs that are distinctive for producing a thick gelatinous capsule that binds cells together into a complex matrix with branching, fingerlike projections. This gelatinous matrix can cause flocculation, the formation of macroscopic particles that settle out of solution. Zoogloea ramigera is of particular importance in aerobic wastewater treatment (Section 22.6), where it degrades much of the organic carbon in the waste stream and promotes flocculation and settling, crucial steps in water purification.
Key Genera: *Chromobacterium, Neisseria*
The order Neisseriales contains at least 29 genera of diverse chemoorganotrophs. The best-characterized species are in the genera Neisseria and Chromobacterium. Species of Neisseria are commonly isolated from animals, and some of them are pathogenic. Neisseria species are always cocci (**Figure 16.9*a***). Some Neisseria are free-living saprophytes and reside in the oral cavity and other moist areas on the animal body. Others are serious pathogens, such as Neisseria meningitidis, which can cause a potentially fatal inflammation of the membranes lining the brain (meningitis, Section 31.5). We discuss the clinical microbiology of Neisseria gonorrhoeae—the causative agent of the sexually transmitted disease gonorrhea—in Section 31.5 and the pathogenesis of gonorrhea itself in Section 31.13.
Figure 16.9 *Neisseria* and *Chromobacterium*.

(a) Transmission electron micrograph of cells of Neisseria gonorrhoeae showing the typical diplococcus cell arrangements. (b) A large colony of Chromobacterium violaceum.
Chromobacterium is a close phylogenetic relative of Neisseria but is rod-shaped in morphology. The best-known Chromobacterium species is C. violaceum, a purple-pigmented organism (Figure 16.9b) found in soil and water and occasionally in pus-forming wounds of humans and other animals. C. violaceum and a few other chromobacteria produce the purple pigment violacein (Figure 16.9b), a water-insoluble pigment with both antimicrobial and antioxidant properties. Chromobacterium is a facultative aerobe, growing fermentatively on sugars and aerobically on various carbon sources.
Key Genera: *Hydrogenophilus, Thiobacillus, Methylophilus, Nitrosomonas*
These three orders contain organisms that have fairly specialized metabolic capabilities including chemolithotrophs and methylotrophs; most species are obligate aerobes and many are autotrophic. Hydrogenophilus thermoluteolus is an obligate aerobe that can grow as a chemolithotroph using H2 as an electron donor for respiration (Section 3.11) and the Calvin cycle to fix CO2. This species is a facultative chemolithotroph but can also grow as a chemoorganotroph on simple carbon sources. Thiobacillus is another important genus of Hydrogenophilales. Species of Thiobacillus can be chemoorganotrophs or chemolithotrophs. Chemolithotrophic species of Thiobacillus are sulfur bacteria (Sections 14.7 and 15.12) that oxidize reduced sulfur compounds as electron donors and grow by aerobic respiration or denitrification (Sections 14.11 and 15.10). Species of Thiobacillus can also fix CO2 using the Calvin cycle and are commonly found in soils, sulfur springs, marine habitats, and other locales where reduced sulfur compounds are available.
The Methylophilales and Nitrosomonadales contain metabolically specialized organisms. Methylophilus species are obligate and facultative methylotrophs (Section 14.16) that grow on methanol and other C1 compounds, but not on CH4. Facultative species can grow as chemoorganotrophs through aerobic respiration of simple sugars. The order Nitrosomonadales contains obligately chemolithotrophic ammonia-oxidizing bacteria, the key genera being Nitrosomonas and Nitrosospira (Section 15.10).
Check Your Understanding
List three species of Betaproteobacteria that are known to be human pathogens.
List three genera of Betaproteobacteria that contain chemolithotrophic species.
Key Genera: *Enterobacter, Escherichia, Klebsiella, Proteus, Salmonella, Serratia, Shigella*
The Gammaproteobacteria are the largest and most diverse class of Proteobacteria, containing nearly half of all characterized species in the phylum. The class contains more than 1500 characterized species and at least 15 well-characterized orders (Figure 16.10, Figure 16.1b). Its species have diverse metabolic and ecological characteristics (Figure 16.2 and Figure 15.1) and include many well-known human pathogens. Species can be phototrophic (including the purple sulfur bacteria, Section 15.4), chemoorganotrophic, or chemolithotrophic, and can have either respiratory or fermentative metabolisms. Members of this group often grow rapidly in laboratory media and can be isolated from a wide diversity of habitats. In this section, we consider the Enterobacteriales, one of the largest and best-known orders within the Gammaproteobacteria.
Figure 16.10 Major orders of *Proteobacteria* in the class *Gammaproteobacteria*.

The phylogenetic tree was constructed using 16S rRNA gene sequences from representative genera of Gammaproteobacteria. Order names are shown in bold.
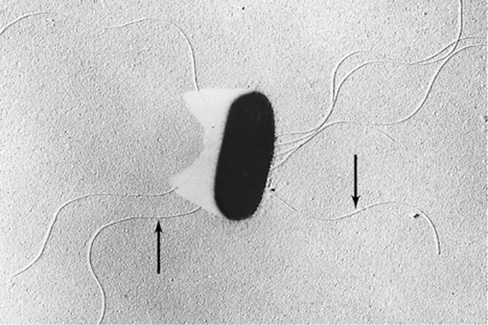
The Enterobacteriales, commonly called the enteric bacteria, comprise a relatively homogeneous phylogenetic group within the Gammaproteobacteria and consist of facultatively aerobic, gram-negative, nonsporulating rods that are either nonmotile or motile by peritrichous flagella (Figure 16.11). The oxidase test and the catalase test are common assays used to characterize bacteria (Section 29.3 and Figure 29.6), and these tests can be used to discriminate enteric bacteria from many other Gammaproteobacteria. The oxidase test is an assay for the presence of cytochrome c oxidase, an enzyme present in many respiring bacteria. The catalase test assays for the enzyme catalase, which detoxifies hydrogen peroxide and is commonly found in bacteria able to grow in the presence of oxygen (Section 4.16 and Figure 4.32). Enteric bacteria are oxidase-negative and catalase-positive. They also produce acid from glucose and reduce nitrate but only to nitrite. Enteric bacteria have relatively simple nutritional requirements and ferment sugars to a variety of end products.
Figure 16.11 A butanediol-producing enteric bacterium.

Electron micrograph of a shadow-cast preparation of a cell of the butanediol-producing bacterium Erwinia carotovora. The cell is about 0.8 μm wide. Note the peritrichously arranged flagella (arrows), typical of enteric bacteria.
Among the enteric bacteria are many species pathogenic to humans, other animals, or plants, as well as other species of industrial importance. Escherichia coli, the best known of all organisms, is the classic enteric bacterium. Because of the medical importance of many enteric bacteria, an extremely large number have been characterized, and numerous genera and species have been defined, largely for ease in identification purposes in clinical microbiology. However, because enteric bacteria are genetically very closely related, their positive identification often presents considerable difficulty. In clinical laboratories, identification is typically based on the combined analysis of a large number of diagnostic tests carried out using miniaturized rapid diagnostic media kits along with immunological and genomic analyses to identify signature proteins or genes of particular species (Chapter 29).
Fermentation Patterns in Enteric Bacteria
One major taxonomic characteristic separating the various genera of enteric bacteria is the type and proportion of fermentation products generated from the fermentation of glucose. Two broad patterns are recognized, the mixed-acid fermentation and the 2,3-butanediol fermentation (Figure 16.12).
Figure 16.12 Enteric fermentations.

Distinction between (a) mixed-acid and (b) butanediol fermentation in enteric bacteria (see also Figure 14.45). The solid arrows indicate reactions leading to major products. Dashed arrows indicate minor products. (a) The photo shows the production of acid (yellow) and gas (in the inverted Durham tube) in a culture of Escherichia coli carrying out a mixed-acid fermentation (purple tube was uninoculated). (b) The photo shows the pink-red color in the Voges–Proskauer (VP) test, which indicates butanediol production, following growth of Enterobacter aerogenes. The left (yellow) tube was not inoculated. Note that the mixed-acid fermentation produces less CO2 but more acid products from glucose than does the butanediol fermentation.
In the mixed-acid fermentation, three acids are formed in significant amounts: acetic, lactic, and succinic. Ethanol, CO2, and H2 are also formed, but not butanediol. In the butanediol fermentation, smaller amounts of acids are formed, and butanediol, ethanol, CO2, and H2 are the main products (Figure 14.45). As a result of mixed-acid fermentation, equal amounts of CO2 and H2 are produced, whereas in the butanediol fermentation, considerably more CO2 than H2 is produced. This is because mixed-acid fermenters produce CO2 only from formic acid by means of the enzyme formate hydrogenlyase: HCOOH→H2+CO2
This reaction results in equal amounts of CO2 and H2. The butanediol fermenters also produce CO2 and H2 from formic acid, but they produce two additional molecules of CO2 during the formation of each molecule of butanediol (Figure 16.12b). Butanediol fermentation is characteristic of Enterobacter, Klebsiella, Erwinia, and Serratia, whereas mixed-acid fermentation is observed in Escherichia, Salmonella, Shigella, Citrobacter, Proteus, and Yersinia.
Mixed-Acid Fermenters: *Escherichia*, *Salmonella*, *Shigella*, and *Proteus*
Species of Escherichia are almost universal inhabitants of the intestinal tract of humans and other warm-blooded animals, although they are by no means the dominant organisms in this habitat. Escherichia may play a nutritional role in the intestinal tract by synthesizing vitamins, particularly vitamin K. As a facultative aerobe, this organism probably also helps consume O2, thus rendering the large intestine anoxic. Wild-type Escherichia strains rarely show any growth-factor requirements and are able to grow on a wide variety of carbon and energy sources such as sugars, amino acids, and organic acids.
Some strains of Escherichia are pathogenic and have been implicated in diarrheal diseases, especially in infants; diarrheal diseases are a major public health problem in developing countries (Sections 33.1, 33.2, 33.7, and 33.11). Escherichia is also a major cause of urinary tract infections in women. Enteropathogenic E. coli strains are becoming more frequently implicated in gastrointestinal infections and generalized fevers. Some strains, such as enterohemorrhagic E. coli, an important representative of which is strain O157:H7, can cause sporadic outbreaks of severe foodborne disease. Infection occurs primarily through consumption of contaminated foods, such as raw or undercooked ground beef, unpasteurized milk, or contaminated water. In a small percentage of cases, E. coli O157:H7 causes a life-threatening complication related to its production of a potent enterotoxin.
Salmonella and Escherichia are quite closely related. However, in contrast to Escherichia, species of Salmonella are almost always pathogenic, either to humans or to other warm-blooded animals (Salmonella is also found in the intestines of cold-blooded animals, such as turtles and lizards). In humans the most common diseases caused by salmonellas are typhoid fever and gastroenteritis (Sections 33.5 and 33.10). The shigellas are also genetically very closely related to Escherichia. Genomic analyses strongly suggest that Shigella and Escherichia have exchanged a significant number of genes by horizontal gene flow. In contrast to most Escherichia, however, species of Shigella are typically pathogenic to humans, causing a rather severe gastroenteritis called bacillary dysentery. Shigella dysenteriae, transmitted by food- and waterborne routes, is a good example of this. The bacterium, which contains endotoxin, invades intestinal epithelial cells, where it excretes a neurotoxin that causes acute gastrointestinal distress.
The genus Proteus typically contains highly motile cells (Figure 16.13) that produce the enzyme urease. Unlike Salmonella and Shigella, Proteus shows only a distant relationship to E. coli. Proteus is a frequent cause of urinary tract infections in humans and probably benefits in this regard from its ready ability to degrade urea by urease. Because of the rapid motility of Proteus cells, colonies growing on agar plates often exhibit a characteristic swarming phenotype (Figure 16.13b). Cells at the edge of the growing colony are more rapidly motile than those in the center of the colony. The former move a short distance away from the colony in a mass and then undergo a reduction in motility, settle down, and divide, forming a new population of motile cells that again swarm. As a result, the mature colony appears as a series of concentric rings, with higher concentrations of cells alternating with lower concentrations (Figure 16.13b).
Figure 16.13 Swarming in *Proteus*.

(a) Cells of Proteus mirabilis stained with a flagella stain; the peritrichous flagella of each cell form into a bundle to rotate in synchrony. (b) Photo of a swarming colony of Proteus vulgaris. Note the concentric rings.
Butanediol Fermenters: *Enterobacter*, *Klebsiella*, and *Serratia*
The butanediol fermenters are genetically more closely related to each other than to the mixed-acid fermenters, a finding that is in agreement with the observed physiological differences (Figure 16.12). Enterobacter aerogenes is a common species in water and sewage as well as the intestinal tract of warm-blooded animals and is an occasional cause of urinary tract infections. One species of Klebsiella, K. pneumoniae, occasionally causes pneumonia in humans, but klebsiellas are most commonly found in soil and water. Most Klebsiella strains also fix nitrogen (Sections 3.12 and 15.9), a property not characteristic of other enteric bacteria.
The genus Serratia forms a series of red pyrrole-containing pigments called prodigiosins (Figure 16.14). Prodigiosin is produced in stationary phase as a secondary metabolite and is of interest because it contains the pyrrole ring also found in the pigments for energy transfer: porphyrins, chlorophylls, bacteriochlorophylls, and phycobilins (Sections 14.3, 14.4 and 14.5). However, it is unclear if prodigiosin plays any role in energy transfer, and its exact function is unknown. Species of Serratia can be isolated from water and soil as well as from the gut of various insects and vertebrates and occasionally from the intestines of humans. Serratia marcescens is also a human pathogen that can cause infections in many body sites. It has been implicated in infections caused by some invasive medical procedures and is an occasional contaminant in intravenous fluids.
Figure 16.14 Colonies of *Serratia marcescens*.
The orange-red pigmentation is due to the pyrrole-containing pigment prodigiosin.
Check Your Understanding
What is a mixed-acid fermentation, and how does it differ from a butanediol fermentation?
What characteristics would you use to distinguish between Escherichia coli and Klebsiella pneumoniae?
16.4 Gammaproteobacteria: Pseudomonadales and Vibrionales
16.4 Gammaproteobacteria: Pseudomonadales and Vibrionales
Key Genera: *Aliivibrio, Pseudomonas, Vibrio*
The phylogenetic and metabolic diversity of the Gammaproteobacteria is remarkable (Figure 16.2 and Figure 15.1), making it difficult to select any particular species as characteristic of the class. We focus here on the Pseudomonadales and Vibrionales, since these groups (along with the Enterobacteriales) represent some of the most commonly encountered orders of Gammaproteobacteria (Figure 16.10).
*Pseudomonadales*
The Pseudomonadales contain exclusively chemoorganotrophs that carry out respiratory metabolisms. All species can grow as aerobes and are typically oxidase- and catalase-positive, but some are also capable of anaerobic respiration with nitrate as the electron acceptor. Most species are able to use a wide diversity of organic compounds as sources of carbon and energy for growth. These organisms are ubiquitous in soil and aquatic systems, and many species cause diseases of plants and animals, including humans. The term pseudomonad is often used to describe any gram-negative, polarly flagellated, aerobic rod that is able to use diverse carbon sources. Pseudomonads can be found in several different groups of Proteobacteria, but here we consider only those organisms in the order Pseudomonadales. The type genus for this order is Pseudomonas.
Several species of Pseudomonas are pathogenic. Among these, Pseudomonas aeruginosa (Figure 16.15) is frequently associated with infections of the urinary and respiratory tracts in humans. P. aeruginosa is not an obligate pathogen. Instead, the organism is an opportunist, initiating infections in individuals with weakened immune systems. P. aeruginosa is a model organism used to study the development of microbial biofilms (Sections 4.9 and 8.10). Due to its ability to readily colonize surfaces, P. aeruginosa is a common cause of hospital-acquired (nosocomial) infections from catheterizations, tracheostomies, lumbar punctures, and intravenous infusions. P. aeruginosa is also a common pathogen in patients receiving treatment for severe burns or other traumatic skin damage; it often infects patients undergoing prolonged treatment with immunosuppressive agents; and it often colonizes the lungs of people with cystic fibrosis. In addition to localized infections, P. aeruginosa can also cause systemic infections, usually in individuals who have experienced extensive skin damage.
Figure 16.15 Cell morphology of pseudomonads.
Shadow-cast transmission electron micrograph of a Pseudomonas cell. The cell measures about 1 μm in diameter.
P. aeruginosa is naturally resistant to many widely used antibiotics, so treatment of infections is often difficult. Resistance is typically due to a resistance transfer plasmid (R plasmid) (Section 6.2 and Section 28.7), which is a plasmid whose genes encode proteins that detoxify various antibiotics or pump them out of the cell. Resistance to antibiotics is further enhanced by biofilm growth (Section 8.10). Polymyxin, an antibiotic not ordinarily used in human therapy because of its toxicity, is effective against P. aeruginosa and is used in critical medical situations.
Certain species of Pseudomonas, such as Pseudomonas syringae, are well-known plant pathogens (phytopathogens). Phytopathogens frequently inhabit nonhost plants (in which disease symptoms are inapparent) and from there become transmitted to host plants and initiate infection. Disease symptoms vary considerably, depending on the particular phytopathogen and host plant. The pathogen releases plant toxins, lytic enzymes, plant growth factors, and other substances that destroy or distort plant tissue, releasing nutrients for use by the bacterium. In many cases the disease symptoms help identify the phytopathogen. Thus, Pseudomonas syringae is typically isolated from leaves showing chlorotic (yellowing) lesions, whereas Pseudomonas marginalis, a “soft-rot” pathogen, infects stems and shoots but rarely leaves.
*Vibrionales*
The Vibrionales contain facultatively aerobic rods and curved rods that employ a fermentative metabolism. One key difference between the Vibrio group and enteric bacteria is that Vibrio are oxidase-positive whereas enteric bacteria are oxidase-negative. Although Pseudomonas species are also oxidase-positive, they are not fermentative and so are clearly distinct from Vibrio species. The best-known genera in this group are Vibrio, Aliivibrio, and Photobacterium, which contain several species that are bioluminescent (Section 23.10).
Most vibrios and related bacteria are aquatic, being found in marine, brackish, or freshwater habitats. Vibrio cholerae is the cause of the disease cholera in humans (Sections 30.8 and 33.3); the organism does not normally cause disease in other hosts. Cholera is one of the most common human infectious diseases in developing countries and is transmitted almost exclusively via water.
Vibrio parahaemolyticus inhabits the marine environment and is a major cause of gastroenteritis in Japan, where raw fish is widely consumed; the organism has also been implicated in outbreaks of gastroenteritis in other parts of the world, including the United States. V. parahaemolyticus can be isolated from seawater itself or from shellfish and crustaceans, and its primary habitat is probably marine animals, with humans being an accidental host.
Check Your Understanding
What species of Pseudomonas is a common cause of lung infection in cystic fibrosis patients? What properties of this species contribute to its pathogenesis?
What major characteristic could be used to differentiate strains of Pseudomonas from those of Vibrio?
16.5 Deltaproteobacteria and Epsilonproteobacteria
16.5 Deltaproteobacteria and Epsilonproteobacteria
16.5 *Deltaproteobacteria* and *Epsilonproteobacteria*
These classes of Proteobacteria contain fewer species and less functional diversity than we have encountered in the Alpha-, Beta-, and Gammaproteobacteria (Figure 16.2 and Figure 15.1). The Deltaproteobacteria are primarily sulfate- and sulfur-reducing bacteria (Sections 14.12, 15.11), dissimilative iron-reducers (Section 15.13), and bacterial predators (Section 15.16). Epsilonproteobacteria, by contrast, contain many species that oxidize the H2S produced by the sulfate and sulfur reducers (Sections 14.7, 15.12). The final class of Proteobacteria, the Zetaproteobacteria, contains only one characterized species (the iron oxidizer Mariprofundus ferrooxydans)and was considered earlier (Section 15.14).
Key Genera: *Bdellovibrio, Myxococcus, Desulfovibrio, Geobacter, Syntrophobacter*
Eight orders have been well characterized within the Deltaproteobacteria (Figure 16.16). The Myxococcales and Bdellovibrionales contain notable genera of bacterial predators (Section 15.16). In contrast, the Desulfuromonadales contains diverse species of metal- and sulfur-reducing genera such as Geobacter (Sections 14.13, 15.13). Indeed, like the Desulfuromonadales, many genera from the Deltaproteobacteria are associated with the reduction of sulfur compounds.
Figure 16.16 Major orders of *Proteobacteria* in the classes *Deltaproteobacteria* and *Epsilonproteobacteria*.

The phylogenetic tree was constructed using 16S rRNA gene sequences from representative genera in the Delta- and Epsilonproteobacteria. Order names are shown in bold.
The largest and most common order containing sulfate reducers is the Desulfovibrionales. These organisms are readily cultivated from marine sediments and nutrient-rich anoxic environments that contain sulfate. Species of Desulfovibrionales are typically incomplete oxidizers (Section 15.11). All use sulfate as the terminal electron acceptor, and all require small organic compounds such as lactate as a source of carbon and energy for growth. Species within the orders Desulfobacterales and Desulfarculales also typically reduce sulfate; however, in contrast to the Desulfovibrionales, these species can be complete or incomplete acetate oxidizers (Section 15.11). In addition to sulfate, some species in these three orders can also reduce sulfite, thiosulfate, or nitrate, and some are capable of certain fermentations.
The final order containing sulfate reducers is the Syntrophobacterales. Some but not all species of the Syntrophobacterales are able to reduce sulfate. In nature, however, species of Syntrophobacterales primarily interact with H2-consuming bacteria in a metabolic partnership called syntrophy (Section 14.22). For example, syntrophic species such as Syntrophobacter wolinii oxidize propionate, producing acetate, CO2, and H2. However, such growth is only possible when a H2-consuming partner is present. If sulfate is present, S. wolinii can grow as a sulfate reducer without the need for a partner. S. wolinii can also grow without a partner organism by fermenting pyruvate, fumarate, or malate.
Key Genera: *Campylobacter, Helicobacter*
The Epsilonproteobacteria (Figure 16.16) were initially defined by pathogenic species of the genera Campylobacter and Helicobacter. While these pathogenic species remain the best characterized in the phylum, environmental studies of marine and terrestrial microbial habitats have shown that a diversity of Epsilonproteobacteria exist in nature. Nearly all of these environmental Epsilonproteobacteria metabolize sulfur compounds in one way or another, and chemolithotrophs and autotrophs are common. We will see that many Epsilonproteobacteria are associated with hosts as either pathogens or symbionts.
*Campylobacter* and *Helicobacter*
These two genera of Epsilonproteobacteria share a number of characteristics. Campylobacter and Helicobacter species are gram-negative, oxidase- and catalase-positive, motile spirilla, and most species are pathogenic to humans or other animals. These organisms are also microaerophilic (Section 4.16) and must therefore be cultured from clinical specimens at low (3–15%) O2 and high (3–10%) CO2.
Campylobacter species, over a dozen of which have been described, cause acute gastroenteritis that typically results in a bloody diarrhea. Pathogenesis is due to several factors, including an enterotoxin that is related to cholera toxin. Helicobacter pylori, also a pathogen, causes both chronic and acute gastritis, leading to the formation of peptic ulcers.We consider these diseases, including their modes of transmission and clinical symptoms, in more detail in Section 31.10.
Sulfur-Metabolizing *Epsilonproteobacteria*
Through environmental sequencing studies (Sections 19.6 and 19.8) and ongoing cultivation efforts, species of Epsilonproteobacteria are now recognized as ubiquitous in marine and terrestrial environments. These microbes are particularly abundant where sulfur-cycling activities are ongoing, particularly in deep-sea hydrothermal vent habitats and marine sediments where sulfide-rich and oxygenated waters mix (Sections 20.15 and 20.16). Thermophilic species are common in this class of bacteria, contributing to their prevalence at hydrothermal systems (Section 20.16). Chemolithotrophy and autotrophy (by the reverse TCA cycle, Section 14.2) are also widespread among Epsilonproteobacteria. Many species are able to grow aerobically or anaerobically (using oxidized nitrogen or sulfur compounds as electron acceptor) while using inorganic electron donors such as reduced sulfur compounds or H2. Epsilonproteobacteria are especially abundant attached to surfaces at the oxic–anoxic interfaces in sulfur-rich environments and play major roles in the oxidation of sulfur compounds in nature.
The ability of Epsilonproteobacteria to fix CO2 is of particular importance to many animals that live in sulfur-rich environments. For example, Epsilonproteobacteria can account for up to 85% of the microbial biomass growing on hydrothermal vent chimneys (Section 20.16 and Figure 20.44). In addition, Epsilonproteobacteria grow as ectosymbionts and endosymbionts of many animals such as oligochaete and polychaete worms, snails, and even shrimp. These symbionts can play two roles for the host, both providing a source of nutrition and helping to detoxify H2S that would otherwise be deleterious to the hosts (Section 23.11). Further exploration of the phylogeny, metabolic activities, and ecological roles of Epsilonproteobacteria will likely uncover exciting new aspects of prokaryotic diversity.
Check Your Understanding
What four metabolic traits are most common in species of Deltaproteobacteria?
What are some characteristic metabolic traits you would expect to find in Epsilonproteobacteria living at a hydrothermal vent?
II *Firmicutes*, *Tenericutes*, and *Actinobacteria*
**The Firmicutes and Actinobacteria contain gram-positive Bacteria and include many well-characterized bacteria. The Tenericutes include species such as Mycoplasma that have lost the ability to make peptidoglycan and a cell wall of any kind.**
We continue our tour of phylogenetic bacterial diversity with the gram-positive bacteria of the phyla Actinobacteria and Firmicutes, and the closely related phylum Tenericutes (Figure 16.17). These three phyla contain nearly half of all characterized species of Bacteria (Figure 16.1b).
Figure 16.17 Major orders of gram-positive bacteria and relatives.

The phylogenetic tree was constructed from 16S rRNA gene sequences of representative genera of Actinobacteria, Firmicutes, and Tenericutes. Order names are shown in bold.
The Actinobacteria include the actinomycetes, a huge group of primarily filamentous soil bacteria. One distinguishing feature of the Actinobacteria is that their genomic DNA typically has a high frequency of GC base pairs, and as a result they are also called the high G + C gram-positive bacteria. The Tenericutes include cells that lack a cell wall, and the Firmicutes include the endospore-forming bacteria, lactic acid bacteria, and several other groups. In contrast to the Actinobacteria, the genomes of Firmicutes generally have a low GC content, and as a result, they are also called the low G + C gram-positive bacteria.
We begin by examining Firmicutes that do not form endospores.
Key Genera: *Lactobacillus, Streptococcus*
The order Lactobacillales contains the lactic acid bacteria, fermentative organisms that produce lactic acid as a major end product of metabolism. These organisms are used widely in food production and preservation (Section 1.6). Lactic acid bacteria are nonsporulating, oxidase- and catalase-negative rods or cocci that show an exclusively fermentative metabolism. All lactic acid bacteria produce lactic acid as a major or sole fermentation product (Sections 3.7, 14.18). Members of this group lack porphyrins and cytochromes; thus they do not carry out oxidative phosphorylation and obtain energy only by substrate-level phosphorylation. However, unlike many anaerobes, most lactic acid bacteria are not sensitive to oxygen (O2) and can grow in its presence; thus they are called aerotolerant anaerobes (Section 4.16).
Most lactic acid bacteria obtain energy only from the metabolism of sugars and therefore are usually restricted to habitats in which sugars are present. They typically have limited biosynthetic abilities, and their complex nutritional requirements include needs for amino acids, vitamins, purines, and pyrimidines (for example, Table 4.2 for Leuconostoc mesenteroides). One important difference between subgroups of the lactic acid bacteria lies in the pattern of products formed from the fermentation of sugars. One group, called homofermentative, produces a single fermentation product, lactic acid. The other group, called heterofermentative, produces other products, mainly ethanol and CO2, as well as lactate (Section 14.18 and Figure 14.44).
Mastering Microbiology
Art Activity: Figure 16.17 Major orders of gram-positive bacteria and relatives
*Lactobacillus*
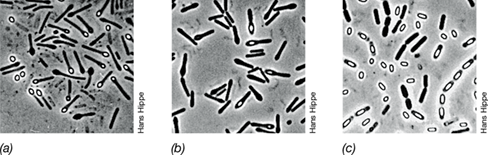
Lactobacilli are typically rod-shaped and grow in chains, varying from long and slender to short, bent rods (Figure 16.18), and most are homofermentative. Lactobacilli are common in dairy products, and some strains are used in the preparation of fermented milk products. For instance, Lactobacillus acidophilus (Figure 16.18a) is used in the production of acidophilus milk; Lactobacillus delbrueckii (Figure 16.18c) is used in the preparation of yogurt; and other species are used in the production of sauerkraut, silage, and pickles (Section 33.6).
Figure 16.18 *Lactobacillus* species.

(a) Lactobacillus acidophilus, phase-contrast. Cells are about 0.75 μm wide. (b) Lactobacillus brevis, transmission electron micrograph. Cells measure about 0.8×2 μm. (c) Lactobacillus delbrueckii, scanning electron micrograph. Cells are about 0.7 μm in diameter.
Lactobacilli are typically more resistant to acidic conditions than are other lactic acid bacteria and are able to grow well at pH values as low as 4. Because of this, they can be selectively enriched from dairy products and fermenting plant material on acidic carbohydrate-containing media. The acid resistance of the lactobacilli enables them to continue growing during natural lactic fermentations, even when the pH value has dropped too low for other lactic acid bacteria to grow. The lactobacilli are therefore typically responsible for the final stages of most lactic acid fermentations. They are rarely, if ever, pathogenic.
*Streptococcus* and Other Cocci
The genera Lactococcus and Streptococcus (Figure 16.19) contain homofermentative species of coccoid-shaped lactic acid bacteria with quite distinct habitats and activities that are of considerable practical importance to humans. Some species are pathogenic to humans and animals (Section 31.2). Streptococcus species (Figure 16.19a) have a characteristic cell morphology of cocci in chains or tetrads and so are readily resolved from the rod-shaped lactobacilli. As producers of lactic acid, other streptococci play important roles in the production of buttermilk, silage, and other fermented products (Section 33.6), and certain species play a major role in the formation of dental caries (Sections 24.3 and 25.1).
Figure 16.19 Gram-positive cocci.

(a) Streptococcus sp., scanning electron micrograph. (b) Lactococcus lactis, phase-contrast micrograph. Cells in both photos are 0.5–1 μm in diameter. (c) Colony of Leuconostoc mesenteroides showing the extensive dextran slime produced by cells grown on sucrose.
There are several other genera of homofermentative cocci. The genus Lactococcus (Figure 16.19b) contains those streptococci of dairy significance, whereas the genus Enterococcus includes streptococci that are primarily of fecal origin and can be human pathogens. Species of the genera Peptococcus and Peptostreptococcus are obligate anaerobes that ferment proteins rather than sugars.
Streptococci have been divided into two groups of related species: the pyogenes subgroup, characterized by Streptococcus pyogenes, the cause of strep throat (Section 31.2), and the viridans subgroup, characterized by Streptococcus mutans, the major cause of dental caries (Sections 24.3 and 25.1). Hemolysis on blood agar is of considerable importance in the subdivision of the genus into species. For example, species that produce the virulence factors streptolysin O or S form colonies surrounded by a large zone of complete red blood cell hemolysis when plated on blood agar, a condition called β-hemolysis (Figures 25.17a, 31.4b, and 31.8). β-hemolysis is diagnostic for streptococci in the pyogenes subgroup. In contrast, streptococci in the viridans subgroup cause incomplete hemolysis on blood agar, a condition that leads to greening of the agar under colonies. Streptococci are also divided into immunological groups (designated by the letters A, B, C, F, G), based on the presence of specific carbohydrate antigens (antigens are substances that elicit an immune response). Those β-hemolytic streptococci found in humans usually contain the group A antigen, whereas enterococci contain the group D antigen.
Heterofermentative lactococci reside in the genus Leuconostoc. Strains of Leuconostoc also produce the flavoring ingredients diacetyl and acetoin from the catabolism of citrate; they have been used as starter cultures in dairy fermentations. Some strains of Leuconostoc produce large amounts of glucose or fructose polysaccharide slimes, especially when cultured on sucrose as the carbon and energy source (Figure 16.19c), and some of these polymers have found medical use as plasma extenders in blood transfusions.
Check Your Understanding
How do heterofermentative and homofermentative bacteria differ physiologically?
How can Streptococcus pyogenes be distinguished from Streptococcus mutans?
16.7 Firmicutes: Nonsporulating Bacillales and Clostridiales
16.7 Firmicutes: Nonsporulating Bacillales and Clostridiales
*Listeria*
The order Bacillales typically contains aerobic and facultatively aerobic chemoorganotrophs. Members of this group are widespread and particularly common in soils. For example, Listeria is found widely in soils and is an opportunistic pathogen and a common cause of foodborne illness. Listeria are gram-positive, catalase-positive, rod-shaped, facultatively aerobic chemoorganotrophs. Although several species of Listeria are known, the species Listeria monocytogenes is most noteworthy because it causes a major foodborne illness, listeriosis (Section 33.13). The organism is transmitted in contaminated, usually ready-to-eat foods such as cheese and sausages and can cause anything from a mild illness to a fatal form of meningitis. Species of Listeria often grow well at low temperatures, allowing growth in refrigerated foods.
Figure 16.20 *Staphylococcus*.

(a) Scanning electron micrograph of typical Staphylococcus aureus cells, showing the irregular arrangement of the cell clusters. Individual cells are about 0.8 μm in diameter. (b) Transmission electron micrograph of a dividing cell of S. aureus. Note the thick gram-positive cell wall.
Staphylococci are common commensals and parasites of humans and animals, and they occasionally cause serious infections. In humans, there are two major species, Staphylococcus epidermidis, a nonpigmented, nonpathogenic organism usually found on the skin or mucous membranes, and Staphylococcus aureus (Figure 16.20), a yellow-pigmented species that is most commonly associated with pathological conditions including boils, pimples, pneumonia, osteomyelitis, meningitis, and arthritis. Some S. aureus strains are resistant to multiple antibiotics (so-called MRSA strains) and are fierce pathogens that can cause extensive tissue damage (Figure 31.9). We discuss the pathogenesis of MRSA and other strains of S. aureus and staphylococcal diseases in Sections 24.5, 29.2, and 31.9.
*Sarcina*
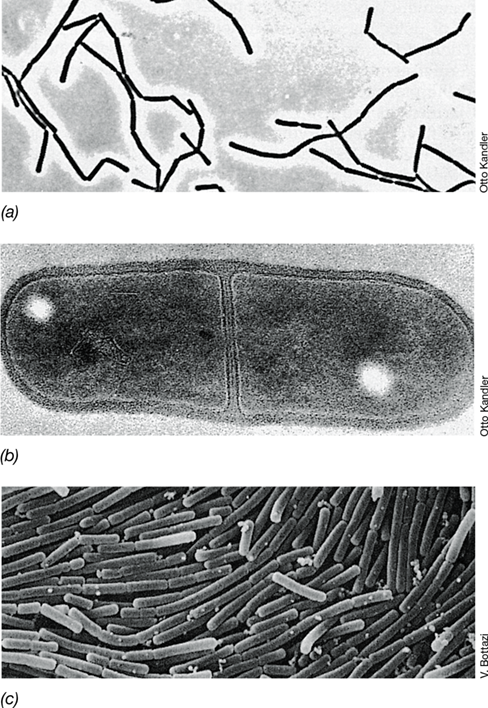
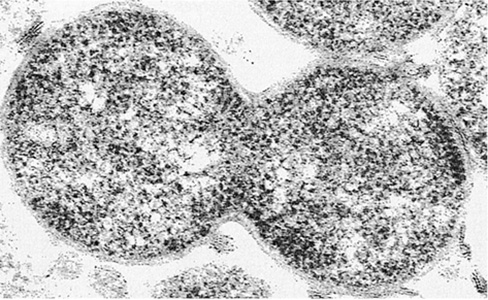
The genus Sarcina contains obligate anaerobes that are catalase-negative within the order Clostridiales. Sarcina species divide in three perpendicular planes to yield packets of eight or more cells and are notable for this morphology (Figure 16.21). Sarcina are also extremely acid-tolerant, being able to ferment sugars and grow in environments at a pH as low as 2. Cells of one species, Sarcina ventriculi, contain a thick, fibrous layer of cellulose surrounding the cell wall (Figure 16.21b). The cellulose layers of adjacent cells become attached, and this functions as a cementing material to hold together packets of S. ventriculi cells.
Figure 16.21 *Sarcina*.

(a) Phase-contrast photomicrograph of cells of a typical gram-positive coccus Sarcina. A single cell is about 2 μm in diameter. (b) Electron micrograph of a thin section from Sarcina ventriculi. The outermost layer of the cell consists of cellulose.
Sarcina species can be isolated from soil, mud, feces, and stomach contents. Because of its extreme acid tolerance, S. ventriculi is one of only a few bacteria that can inhabit and grow in the stomach of humans and other monogastric animals. Rapid growth of S. ventriculi is observed in the stomach of humans suffering from certain gastrointestinal disorders, such as pyloric ulcerations. These pathological conditions retard the flow of food to the intestine and often require surgery to correct.
Check Your Understanding
How could species of Staphylococcus be differentiated from Streptococcus?
What characteristics differentiate Sarcina from Staphylococcus?
16.8 Firmicutes: Sporulating Bacillales and Clostridiales
16.8 Firmicutes: Sporulating Bacillales and Clostridiales
Key Genera: *Bacillus, Clostridium, Sporosarcina*
All endospore-forming bacteria are gram-positive species of Bacillales or Clostridiales. The ability to form endospores evolved only once in a common ancestor of the Bacillales, Clostridiales, and Lactobacillales (Figure 16.17). However, many Bacillales and Clostridiales and the entire order Lactobacillales are unable to form endospores. The capacity to make endospores requires many genes (Sections 2.8 and 8.6) and has not been acquired by horizontal gene transfer. It thus appears that the phylogenetic distribution of endospores has seen many cases where the capacity to form endospores has been lost during the course of evolution.
Endospore-forming bacteria are distinguished on the basis of cell morphology, shape and cellular position of the endospore (Figure 16.22), relationship to O2, and energy metabolism. The two genera about which most is known are Bacillus, species of which are aerobic or facultatively aerobic, and Clostridium, which contains species that are obligately anaerobic and fermentative. All endospore-forming bacteria are ecologically related because they are found in nature primarily in soil. Even those species that are pathogenic to humans or other animals are primarily saprophytic soil organisms and infect animals only incidentally. Indeed, the ability to produce endospores should be advantageous for a soil microorganism because soil is a highly variable environment in terms of nutrient levels, temperature, and water activity.
Figure 16.22 *Clostridium* species and endospore location.

(a) Clostridium cadaveris, terminal endospores. Cells are about 0.9 μm wide. (b) Clostridium sporogenes, subterminal endospores. Cells are about 1 μm wide. (c) Clostridium bifermentans, central endospores. Cells are about 1.2 μm wide. All are phase-contrast micrographs.
Endospore-forming bacteria can be selectively isolated from soil, food, dust, and other materials by heating the sample to 80 °C for 10 min, a treatment that effectively kills vegetative cells while any endospores present remain viable. Streaking such heat-treated samples on plates of the appropriate medium and incubating either aerobically or anaerobically selectively yields species of Bacillus or Clostridium, respectively.
*Bacillus* and *Paenibacillus*
Species of Bacillus and Paenibacillus grow well on defined media containing any of a number of carbon sources. Many bacilli produce extracellular hydrolytic enzymes that break down complex polymers such as polysaccharides, nucleic acids, and lipids, permitting the organisms to use these products as carbon sources and electron donors. Many bacilli produce antibiotics, including bacitracin, polymyxin, tyrocidine, gramicidin, and circulin. In most cases the antibiotics are released when the culture enters the stationary phase of growth and is committed to sporulation.
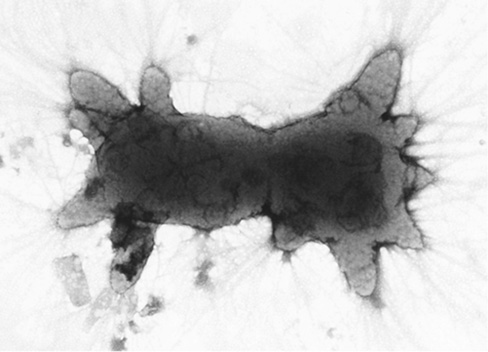
Several bacilli, most notably Paenibacillus popilliae and Bacillus thuringiensis, produce toxic insecticidal proteins. P. popilliae causes a fatal condition called milky disease in Japanese beetle larvae and larvae of closely related beetles of the family Scarabaeidae. B. thuringiensis causes a fatal disease of many different groups of insects. Both of these insect pathogens form a crystalline protein during sporulation called the parasporal body, which is deposited within the sporangium but outside the endospore proper (Figure 16.23). In B. thuringiensis, the parasporal body is a protoxin that is converted to a toxin in the insect gut. The toxin binds to specific receptors in the intestinal epithelial cells of certain insects and induces pore formation that causes leakage of the host cell cytoplasm followed by lysis. Diverse strains of B. thuringiensis can make different types of toxin that have specificity for different groups of insects. Endospore preparations derived from B. thuringiensis and P. popilliae are commercially available as biological insecticides.
Figure 16.23 The toxic parasporal crystal in the insect pathogen *Bacillus thuringiensis*.

Electron micrograph of a thin section of a sporulating cell. The crystalline protein (Bt toxin) is toxic to certain insects by causing lysis of their intestinal cells.
The cry genes that encode crystal proteins have been isolated from several B. thuringiensis strains. The genes for the B. thuringiensis crystal protein (known commercially as “Bt toxin”) have been introduced into genetically modified crops (e.g., maize, soybeans, and cotton) to render the plants resistant to insects. These genetically modified “Bt crops” are used widely around the world.
*Clostridium*
Clostridia lack a respiratory chain, unlike Bacillus species, and so they obtain ATP by substrate-level phosphorylation. Many anaerobic energy-yielding mechanisms are known in the clostridia (Section 14.19). Indeed, the separation of the genus Clostridium into subgroups is based primarily on these properties and on the fermentable substrate used. A number of clostridia are saccharolytic and ferment sugars, producing butyric acid as a major end product. Some of these also produce acetone and butanol, such as Clostridium pasteurianum, which is also a vigorous nitrogen-fixing bacterium (Sections 3.12 and 15.9).
One group of clostridia including the species C. thermocellum, C. cellulolyticum, and C. cellulovorans ferments cellulose with the formation of acids and alcohols. These species are likely the major organisms decomposing cellulose in anoxic environments such as the rumen and sediments. Cellulolytic clostridia possess cellulosomes, complex multienzyme structures found on the outer surface of the cell wall. The cellulosome binds insoluble cellulose and degrades it into soluble products that are transported into the cytoplasm and metabolized by the cell. This cellulosome mechanism is common to bacteria that are able to degrade cellulose anaerobically.
Another group of clostridia are proteolytic and conserve energy from the fermentation of amino acids. Some species ferment individual amino acids, but others ferment only amino acid pairs. The products of amino acid fermentation are typically acetate, butyrate, CO2, and H2. The coupled catabolism of an amino acid pair is called a Stickland reaction; for example, Clostridium sporogenes ferments glycine plus alanine. In the Stickland reaction, one amino acid functions as the electron donor and is oxidized, whereas the other is the electron acceptor and is reduced (Figure 14.47). Many of the products of amino acid fermentation by clostridia are foul-smelling substances, and the odor that results from putrefaction is mainly the result of clostridial action. In addition to butyric acid, other odoriferous compounds produced are isobutyric acid, isovaleric acid, caproic acid, hydrogen sulfide, methyl mercaptan (from sulfur amino acids), cadaverine (from lysine), putrescine (from ornithine), and ammonia.
The main habitat of clostridia is the soil, where they live primarily in anoxic “pockets” made free of O2 by the respiratory activities of facultative or obligately aerobic bacteria. In addition, a number of clostridia inhabit the anoxic environment of the mammalian intestinal tract. Several clostridia are capable of causing severe diseases in humans, as will be discussed in Sections 24.9, 30.7, 32.9 and 33.9. For example, botulism is caused by Clostridium botulinum, tetanus by Clostridium tetani, and gas gangrene by Clostridium perfringens and a number of other clostridia, both sugar and amino acid fermenters. These pathogenic clostridia seem in no way unusual metabolically but are distinct in that they produce specific toxins or, in those causing gas gangrene, a group of toxins. C. perfringens and related species can also cause gastroenteritis in humans and domestic animals (Section 33.9), and botulism outbreaks are not uncommon in birds such as ducks and a variety of other animals.
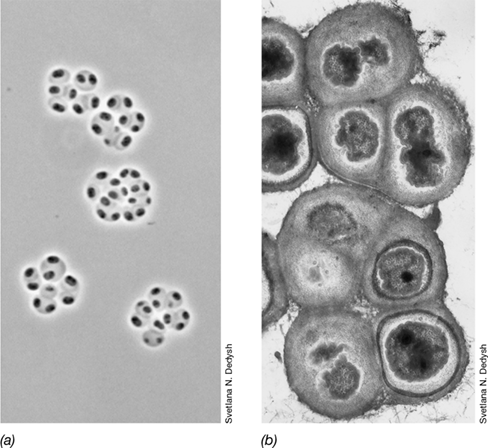
*Sporosarcina*
The genus Sporosarcina (Figure 16.24) is unusual among endospore formers because cells are cocci instead of rods. Sporosarcina consists of strictly aerobic spherical to oval cells that divide in two or three perpendicular planes to form tetrads or packets of eight or more cells. The major species is Sporosarcina ureae. This bacterium can be enriched from soil by plating dilutions of a pasteurized soil sample on alkaline nutrient agar supplemented with 8% urea and incubating in air. Most soil bacteria are strongly inhibited by as little as 2% urea. However, S. ureae tolerates this, catabolizing urea to CO2 and ammonia (NH3), which dramatically raises the pH. S. ureae is remarkably alkaline-tolerant and can be grown in media up to pH 10, and this feature can be used to advantage in its enrichment from soil.
Figure 16.24 *Sporosarcina ureae.*

Phase-contrast micrograph. A single cell is about 2 μm wide. Note bright refractile endospores. Most cell packets contain eight cells.
Check Your Understanding
What is the major physiological distinction between Bacillus and Clostridium species?
What is the crystalline protein made by Bacillus thuringiensis and what is its significance to agriculture?
Key Genera: *Mycoplasma, Spiroplasma*
The Tenericutes, which contain the single class Mollicutes, are bacteria that lack cell walls (mollis is Latin for “soft”) and are some of the smallest organisms known. This group is often called the mycoplasmas because Mycoplasma, a notable genus containing human pathogens, is the best-characterized genus in the phylum.
Although they do not stain gram-positively (because they lack cell walls), mycoplasmas are phylogenetically related to the Firmicutes. Mycoplasmas typically live within animal and plant hosts and this may eliminate the need for a gram-positive cell wall. These organisms also have small genomes (ranging in size from 0.6 to 2.2 megabase pairs [Mbp]), a characteristic common in obligate symbionts and intracellular pathogens (Section 10.3 and Section 23.7).
Properties of Mycoplasmas
The absence of cell walls in mycoplasmas has been confirmed by electron microscopy and chemical analyses, which show that peptidoglycan is absent. Mycoplasmas resemble protoplasts (bacteria treated to remove their cell walls), but they are more resistant to osmotic lysis and are able to survive conditions under which protoplasts lyse. This ability to resist osmotic lysis is at least partially determined by the presence of sterols, which make the cytoplasmic membrane of mycoplasmas more stable than that of other bacteria. Indeed, some mycoplasmas require sterols in their growth media, and this sterol requirement can aid in the classification of mycoplasmas.
In addition to sterols, certain mycoplasmas contain compounds called lipoglycans. Lipoglycans are long-chain heteropolysaccharides covalently linked to membrane lipids and embedded in the cytoplasmic membrane of many mycoplasmas. Lipoglycans in some ways resemble the lipopolysaccharides in the outer membrane of gram-negative bacteria, except that they lack the lipid A backbone (Section 2.4). Lipoglycans function to help stabilize the cytoplasmic membrane and have also been identified as facilitating attachment of mycoplasmas to cell surface receptors of animal cells.
Growth of Mycoplasmas
Mycoplasmas can be grown in the laboratory and are small and pleomorphic cells. A single culture may exhibit small coccoid elements; larger, swollen forms; and filamentous forms, often highly branched (Figure 16.25). The small coccoid elements (0.2–0.3 μm in size) are among the smallest of free-living cells. The mode of growth of mycoplasmas differs in liquid and agar cultures. On agar the organisms tend to grow so that they become embedded in the medium. These colonies show a characteristic “fried-egg” appearance consisting of a dense central core that penetrates downward into the agar, surrounded by a circular spreading area that is lighter in color (Figure 16.26). As would be expected of cells lacking cell walls, growth of Mollicutes is not inhibited by antibiotics that inhibit cell wall synthesis. However, mycoplasmas are as sensitive as most Bacteria to antibiotics whose targets are other than the cell wall.
Figure 16.25 *Mycoplasma mycoides*.
Metal-shadowed transmission electron micrograph. Note the coccoid and hyphae-like elements. The average diameter of cells in chains is about 0.5 μm.
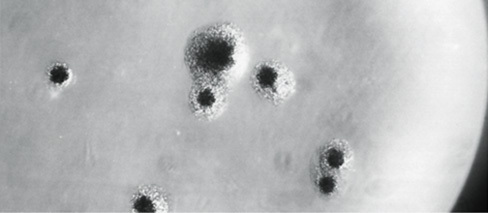
Figure 16.26 Colonies of a *Mycoplasma* species on agar.
Note the typical “fried-egg” appearance. The colonies are about 0.5 mm in diameter.
Media for the culture of mycoplasmas are typically quite complex. For many species, growth is poor or absent even in complex yeast extract–peptone–beef heart infusion media. Fresh serum or ascitic fluid (peritoneal fluid) is needed as well to provide unsaturated fatty acids and sterols. Some mycoplasmas can be cultivated on relatively simple culture media, however, and even defined media have been developed for some species. Most mycoplasmas use carbohydrates as carbon and energy sources and require vitamins, amino acids, purines, and pyrimidines as growth factors.
*Spiroplasma*
The genus Spiroplasma consists of helical or spiral-shaped Mollicutes. Amazingly, although they lack a cell wall and flagella, spiroplasmas are motile by means of a rotary (screw) motion or a slow undulation. Intracellular fibrils that are thought to play a role in motility have been demonstrated. The organism has been isolated from ticks, the hemolymph (Figure 16.27) and gut of insects, vascular plant fluids and insects that feed on these fluids, and the surfaces of flowers and other plant parts. For example, Spiroplasma citri has been isolated from the leaves of citrus plants, where it causes a disease called citrus stubborn disease, and from corn plants suffering from corn stunt disease. A number of other mycoplasma-like organisms have been detected in diseased plants by electron microscopy, which indicates that a large group of plant-associated Mollicutes may exist. Some species of Spiroplasma are known that cause insect diseases, such as honeybee spiroplasmosis and lethargy disease of the beetle Melolontha.
Figure 16.27 “Sex-ratio” spiroplasma from the hemolymph of the fly *Drosophila pseudoobscura*.
Dark-field micrograph. Female flies infected with the sex-ratio spiroplasma bear only female progeny. Cells are about 0.15 μm in diameter.
Check Your Understanding
Why do mycoplasmas need to have stronger cytoplasmic membranes than other bacteria?
Motile spiroplasmas cannot contain a normal bacterial flagellum; why?
16.10 Actinobacteria: Coryneform and Propionic Acid Bacteria
16.10 Actinobacteria: Coryneform and Propionic Acid Bacteria
Key Genera: *Arthrobacter, Corynebacterium, Propionibacterium*
The other major group of gram-positive bacteria is the Actinobacteria, which form their own phylum within the Bacteria. The Actinobacteria contain rod-shaped to filamentous and primarily aerobic bacteria that are common inhabitants of soil and plant materials. For the most part they are harmless commensals, species of Mycobacterium (for example, Mycobacterium tuberculosis) being notable exceptions. Some are of great economic value in either the production of antibiotics or certain fermented dairy products. While there are nine orders of Actinobacteria, the vast majority of species belong to the order Actinomycetales (Figure 16.17). We consider here the coryneform bacteria, species of Actinomycetales that have an unusual method of cell division, and the propionic acid bacteria, important agents in the ripening of Swiss cheese.
Coryneform Bacteria
Coryneform bacteria are gram-positive, aerobic, nonmotile, rod-shaped organisms that form irregular-shaped, club-shaped, or V-shaped cell arrangements during growth. V-shaped cells arise as a result of an abrupt movement that occurs just after cell division, a process called snapping division (Figure 16.28). Snapping division occurs because the cell wall consists of two layers; only the inner layer participates in cross-wall formation, and so after the cross-wall is formed, the two daughter cells remain attached by the outer layer of the cell wall. Localized rupture of this outer layer on only one side of the cell results in a bending of the two cells away from the ruptured side (Figure 16.29) and thus development of V-shaped forms.
Figure 16.28 Snapping division in *Arthrobacter*.

Phase-contrast micrograph of characteristic V-shaped cell groups in Arthrobacter crystallopoietes resulting from snapping division. Cells are about 0.9 μm in diameter.
Figure 16.29 Cell division in *Arthrobacter*.

Transmission electron micrograph of cell division in Arthrobacter crystallopoietes, illustrating how snapping division and V-shaped cell groups arise. (a) Before rupture of the outer cell wall layer (arrow). (b) After rupture of the outer layer on one side. Cells are 0.9–1 μm in diameter.
The main genera of coryneform bacteria are Corynebacterium and Arthrobacter. The genus Corynebacterium consists of an extremely diverse group of bacteria, including animal and plant pathogens and saprophytes. Some species, such as Corynebacterium diphtheriae, are pathogenic (diphtheria, Section 31.3). The genus Arthrobacter, consisting primarily of soil organisms, is distinguished from Corynebacterium on the basis of a developmental cycle during which it changes morphology between rod and coccus (Figure 16.30). Corynebacterium cells frequently have a swollen end, so they have a club-shaped appearance, whereas Arthrobacter species are less commonly club-shaped.
Figure 16.30 Stages in the life cycle of *Arthrobacter globiformis* as observed in slide culture.

(a) Single coccoid element; (b–e) conversion to rod and growth of a microcolony consisting predominantly of rods; (f–g) conversion of rods to coccoid forms. Cells are about 0.9 μm in diameter.
Along with the Acidobacteria (Section 16.21), species of Arthrobacter are among the most common of all soil bacteria. They are remarkably resistant to desiccation and starvation, despite the fact that they do not form spores or other resting cells. Arthrobacters are a heterogeneous group that have considerable nutritional versatility, and strains have been isolated that decompose herbicides, caffeine, nicotine, phenols, and other unusual organic compounds.
Propionic Acid Bacteria
The propionic acid bacteria (genus Propionibacterium) were first discovered in Swiss (Emmentaler) cheese, where their fermentative production of CO2 produces the characteristic holes. In addition, the propionic acid they produce is at least partly responsible for the unique flavor of the cheese. The bacteria in this group are gram-positive anaerobes that ferment lactic acid, carbohydrates, and polyhydroxy alcohols, producing primarily propionic acid, acetic acid, and CO2 (Section 14.20).
The fermentation of lactate is of interest because lactate itself is a fermentation product of many bacteria (Section 16.6). The starter culture in Swiss cheese manufacture consists of a mixture of homofermentative streptococci and lactobacilli, plus propionic acid bacteria. The homofermentative organisms carry out the initial fermentation of lactose to lactic acid during formation of the curd (protein and fat). After the curd has been drained, the propionic acid bacteria develop rapidly. The eyes (or holes) characteristic of Swiss cheese are formed by the accumulation of CO2, the gas diffusing through the curd and gathering at weak points. The propionic acid bacteria are thus able to obtain energy anaerobically from a product that other bacteria have produced by fermentation. This metabolic strategy is called a secondary fermentation.
Propionate is also formed in the fermentation of succinate by the bacterium Propionigenium. This organism is phylogenetically and ecologically unrelated to Propionibacterium, but energetic aspects of its fermentation are of considerable interest.We discussed the mechanism of the Propionigenium fermentation in Section 14.20.
Check Your Understanding
What is snapping division and what organism exhibits it?
What organism is involved in the production of Swiss cheese, and what products does it make that help to flavor the cheese and make the holes?
Key Genus: *Mycobacterium*
Mycobacteria are common in soils and most are harmless, but the genus Mycobacterium contains several notable human pathogens, chief among them Mycobacterium tuberculosis, the cause of tuberculosis (Section 31.4). Species are rod-shaped bacteria that at some stage of their growth cycle possess the distinctive staining property called acid-fastness. This property is due to the presence of unique lipids called mycolic acids, found only in species of the genus Mycobacterium, on the surface of the mycobacterial cell. Mycolic acids are a group of complex branched-chain hydroxylated lipids (**Figure 16.31*a***) covalently bound to peptidoglycan in the cell wall; the complex gives the cell surface a waxy, hydrophobic consistency.
Figure 16.31 Acid-fast staining.

Structure of (a) mycolic acid and (b) basic fuchsin, the dye used in the acid-fast stain. The fuchsin dye combines with mycolic acids in the cell wall via ionic bonds between COO− and NH2 +. Inset: Acid-fast stain of cells (red) of Mycobacterium tuberculosis present in a sputum sample from a tuberculosis patient.
Because of their waxy surface, mycobacteria do not stain well with Gram stain. A mixture of the red dye basic fuchsin and phenol is used in the acid-fast (Ziehl–Neelsen) stain. The stain is driven into the cells by slow heating, and the role of the phenol is to enhance penetration of the fuchsin into the lipids. After washing in distilled water, the preparation is decolorized with acid alcohol and counterstained with methylene blue. Cells of acid-fast organisms stain red (Figure 16.31 inset), whereas the background and non-acid-fast organisms appear blue (Figure 31.15a).
Mycobacteria are somewhat pleomorphic and may undergo branching or even filamentous growth. However, in contrast to the filaments of the actinomycetes (Section 16.12), the filaments of the mycobacteria do not form a true mycelium. Mycobacteria can be separated into two major groups: slow-growing species (e.g., M. tuberculosis, M. avium, M. bovis, and M. gordonae) and fast-growing species (e.g., M. smegmatis, M. phlei, M. chelonae, M. parafortuitum). Mycobacterium tuberculosis is a typical slow grower, and visible colonies are produced from dilute inoculum only after days to weeks of incubation. When growing on solid media, mycobacteria form tight, compact, often wrinkled colonies (Figure 16.32). This colony morphology is probably due to the high lipid content and hydrophobic nature of the cell surface, which facilitates cells sticking together.
Figure 16.32 Characteristic colony morphology of mycobacteria.

(a) Mycobacterium tuberculosis, showing the compact, wrinkled appearance of the colony. The colony is about 7 mm in diameter. (b) A colony of virulent M. tuberculosis at an early stage, showing the characteristic cordlike growth. Individual cells are about 0.5 μm in diameter. (See also the historic drawings of M. tuberculosis cells made by Robert Koch, Figure 1.35). (c) Colonies of Mycobacterium avium from a strain of this organism isolated as an opportunistic pathogen from an AIDS patient.
For the most part, mycobacteria have relatively simple nutritional requirements. Most species can grow aerobically in a simple mineral-salts medium with ammonium as the nitrogen source and glycerol or acetate as the sole carbon source and electron donor. Growth of M. tuberculosis is more difficult and is stimulated by lipids and fatty acids. The virulence of M. tuberculosis cultures has been correlated with the formation of long, cordlike structures (Figure 16.32b) that form as a result of side-to-side aggregation and intertwining of long chains of bacteria. Growth in cords reflects the presence of a characteristic glycolipid, the cord factor, on the cell surface (Figure 16.33). The pathogenesis of tuberculosis, along with the related mycobacterial disease leprosy, is discussed in Section 31.00.
Figure 16.33 Structure of cord factor, a mycobacterial glycolipid: 6,6′-di-*O*-mycolyl trehalose.

The two identical long-chain dialcohol groups are shown in purple. Inset: Photomicrograph of acid-fast stained cells of Mycobacterium tuberculosis (Figure 16.31) that have formed cords.
Some mycobacteria produce yellow carotenoid pigments (Figure 16.32c), and pigmentation can aid in identification. Mycobacteria can either be nonpigmented (e.g., M. tuberculosis, M. bovis, M. smegmatis, M. chelonae); or can form pigment only when cultured in light, a property called photochromogenesis (e.g., M. parafortuitum); or can form pigment even when cultured in the dark, a property called scotochromogenesis (e.g., M. gordonae, M. phlei). Photochromogenesis is triggered by the blue region of the visible spectrum and is characterized by the photoinduction of one of the early enzymes in carotenoid biosynthesis. As with other carotenoid-containing bacteria, it is likely that carotenoids protect mycobacteria against oxidative damage from singlet oxygen (Section 4.16).
Check Your Understanding
What is mycolic acid, and what properties does this substance confer on mycobacteria?
16.12 Filamentous Actinobacteria: Streptomyces and Relatives
16.12 Filamentous Actinobacteria: Streptomyces and Relatives
Key Genera: *Streptomyces, Actinomyces, Nocardia*
The actinomycetes are a large group of phylogenetically related, filamentous and aerobic gram-positive Bacteria common in soils. Many actinomycetes have a characteristic developmental cycle that culminates in the production of desiccation-resistant spores. Filaments elongate from their ends and form branching hyphae. Hyphal growth results in a network of filaments called a mycelium (Figure 16.34), analogous to that formed by filamentous fungi (Section 18.9). When nutrients are depleted, the mycelium forms aerial hyphae that differentiate into spores that allow for survival and dispersal. We focus here on the genus Streptomyces, the most important genus in this group.
Figure 16.34 *Nocardia*.
A young colony of an actinomycete of the genus Nocardia, showing typical filamentous cellular structure (mycelium). Each filament is about 0.8–1 μm in diameter.
*Streptomyces*
Over 600 species of Streptomyces are recognized. Streptomyces filaments are typically 0.5–1.0 μm in diameter and of indefinite length, and often lack cross-walls in the vegetative phase. Streptomyces grow at the tips of the filaments and may branch often. Thus, the vegetative phase consists of a complex, tightly woven matrix, resulting in a compact, convoluted mycelium and subsequent colony. As the colony ages, characteristic aerial filaments called sporophores are formed, which project above the surface of the colony and give rise to spores (Figure 16.35).
Figure 16.35 Spore-bearing structures of actinomycetes.

Phase-contrast micrographs. Compare these photos with the art in Figure 16.37. (a) Streptomyces, a monoverticillate type. (b) Streptomyces, a closed spiral type. Filaments are about 0.8 μm wide in both types.
Streptomyces spores, called conidia, are quite distinct from the endospores of Bacillus and Clostridium. Unlike the elaborate cellular differentiation that leads to the formation of an endospore, conidia are produced by the formation of cross-walls in the multinucleate sporophores followed by separation of the individual cells directly into spores (Figure 16.36). Differences in the shape and arrangement of aerial filaments and spore-bearing structures of various species are among the fundamental features used in classifying the Streptomyces species (Figure 16.37). The conidia and sporophores are often pigmented and contribute a characteristic color to the mature colony (Figure 16.38). The dusty appearance of the mature colony, its compact nature, and its color make detection of Streptomyces colonies on agar plates relatively easy (Figure 16.38b).
Figure 16.36 Spore formation in *Streptomyces*.

Diagram of stages in the conversion of an aerial hypha (sporophore) into spores (conidia).
Figure 16.37 Morphologies of spore-bearing structures in the streptomycetes.

A given species of Streptomyces produces only one morphological type of spore-bearing structure. The term “verticillate” means “whorls.”
Figure 16.38 Streptomycetes.

(a) Colonies of Streptomyces and other soil bacteria derived from spreading a soil dilution on a casein–starch agar plate. The Streptomyces colonies are of various colors (several black Streptomyces colonies are near the top of the plate) but can easily be identified by their opaque, rough, nonspreading morphology. (b) Close-up photo of colonies of Streptomyces coelicolor.
Mastering Microbiology
Art Activity: Figure 16.36 Spore formation in Streptomyces
Ecology and Isolation of *Streptomyces*
Although a few streptomycetes are aquatic, they are primarily soil organisms. In fact, the characteristic earthy odor of soil is caused by the production by streptomycetes of a series of complex metabolites all called geosmin. Alkaline to neutral soils are more favorable for the development of Streptomyces than are acid soils. Moreover, higher numbers of Streptomyces are found in well-drained soils (such as sandy loams or soils covering limestone), where conditions are more likely to be aerobic, than in waterlogged soils, which quickly become anoxic.
Isolation of Streptomyces from soil is relatively easy: A suspension of soil in sterile water is diluted and spread on selective agar medium, and the plates are incubated aerobically at 25 °C (Figure 16.38). Media selective for Streptomyces contain mineral salts plus polymeric substances such as starch or casein as organic nutrients. Streptomycetes typically produce extracellular hydrolytic enzymes that permit utilization of polysaccharides (starch, cellulose, and hemicellulose), proteins, and fats, and some strains can use hydrocarbons, lignin, tannin, and other polymers. After incubation for 5–7 days in air, the plates are examined for the presence of the characteristic Streptomyces colonies (Figure 16.38), and spores from colonies can be restreaked to isolate pure cultures.
Antibiotics of *Streptomyces*
Perhaps the most striking physiological property of the streptomycetes is the extent to which they produce antibiotics (Table 16.2). Evidence for antibiotic production is often seen on the agar plates used in their initial isolation: Adjacent colonies of other bacteria show zones of inhibition (**Figure 16.39*a***).
Table 16.2 Some common antibiotics synthesized by species of *Streptomyces* and related *Actinobacteria*

a Most antibiotics are effective against several different Bacteria. The entries in this column refer to the common clinical application of a given antibiotic.The structures and mode of action of many of these antibiotics are discussed in Sections 28.5, 28.6 and 28.7.
b All species names beginning with an “S.” are species of Streptomyces.
Figure 16.39 Antibiotics from *Streptomyces*.

(a) Antibiotic action of soil microorganisms on a crowded agar plate. The smaller colonies surrounded by inhibition zones (arrows) are streptomycetes; the larger, spreading colonies are Bacillus species, some of which are also producing antibiotics. (b) The red-colored antibiotic undecylprodigiosin is being excreted by colonies of S. coelicolor.
About 50% of all Streptomyces isolated have been found to be antibiotic producers. Over 500 distinct antibiotics are produced by streptomycetes and many more are suspected; most of these have been identified chemically. Some species produce more than one antibiotic, and often the several antibiotics produced by one organism are chemically unrelated. Although an antibiotic-producing organism is resistant to its own antibiotics, it usually remains sensitive to antibiotics produced by other streptomycetes. Many genes are required to encode the enzymes for antibiotic synthesis, and because of this, the genomes of Streptomyces species are typically quite large (8 Mbp and larger; compare with genome sizes in Table 10.1). More than 60 streptomycete antibiotics have been used in human and veterinary medicine, and some of the most commonly used are listed in Table 16.2.
Surprisingly, despite the extensive research done on antibiotic-producing streptomycetes by the antibiotic industry and the fact that Streptomyces antibiotics are a multibillion-dollar-a-year industry, the ecology of Streptomyces remains poorly understood. The interactions of these organisms with other bacteria and the ecological rationale for antibiotic production remains an important topic about which we know very little. One hypothesis for why Streptomyces species produce antibiotics is that antibiotic production, which is linked to sporulation (a process itself triggered by nutrient depletion), might be a mechanism to inhibit the growth of other organisms competing with Streptomyces cells for limiting nutrients. This would allow the Streptomyces to complete the sporulation process and form a dormant structure that would increase their chances of survival.
Check Your Understanding
Contrast spores and sporulation in Streptomyces and Bacillus species.
Why might antibiotic production be of advantage to streptomycetes?
III *Bacteroidetes*
**The Bacteroidetes include aerobes such as Cytophaga, which are common in natural habitats, and obligate anaerobes such as Bacteroides, which are common in the guts of many animals. Many of these species are motile by gliding.**
The phylum Bacteroidetes contains more than 1000 characterized species spread across four primary orders: Bacteroidales, Cytophagales, Flavobacteriales, and Sphingobacteriales (Figure 16.40). The Bacteroidetes are gram-negative, nonsporulating rods; species are typically saccharolytic and can be aerobic or fermentative, including obligate aerobes, facultative aerobes, and obligate anaerobes. Gliding motility (Section 2.10) is widespread in the phylum, though many species are nonmotile and a few are motile by flagella. The genus Bacteroides has been particularly well studied as these organisms are a major component of the microbial community in the human gut. But as we will see in Chapter 20, as a group, Bacteroidetes are found almost everywhere in nature and are diverse and common inhabitants of most terrestrial and aquatic environments.
Figure 16.40 Major orders of *Bacteroidetes*.

The phylogenetic tree was constructed from 16S rRNA gene sequences of representative genera of Bacteroidetes. Order names are shown in bold.
Key Genus: *Bacteroides*
The order Bacteroidales primarily contains obligately anaerobic fermentative species. The type genus is Bacteroides, which contains species that are saccharolytic, fermenting sugars or proteins (depending on the species) to acetate and succinate as major fermentation products. Bacteroides are normally commensals, found in the intestinal tract of humans and other animals. In fact, Bacteroides species are the numerically dominant bacteria in the human large intestine, where measurements have shown that about 1011 prokaryotic cells are present per gram of wet feces (Section 24.2). However, species of Bacteroides can occasionally be pathogens and are the most important anaerobic bacteria associated with human infections such as bacteremia (bacteria in the blood).
Bacteroides thetaiotaomicron is one of the most prominent species of Bacteroides found in the lumen of the large intestine. B. thetaiotaomicron specializes in the degradation of complex polysaccharides. A majority of its genome is devoted to making enzymes that degrade polysaccharides. The diversity and number of genes for carbohydrate metabolism found in its genome far exceeds those found in any other bacterial species. B. thetaiotaomicron produces many enzymes that are not encoded by the human genome and thus it vastly increases the diversity of plant polymers that can be degraded in the human digestive tract.
Species of Bacteroides are unusual in that they are one of the few groups of bacteria to synthesize a special type of lipid called sphingolipid (Figure 16.41), a collection of lipids characterized by the long-chain amino alcohol sphingosine in place of glycerol in the lipid backbone. Sphingolipids such as sphingomyelin, cerebrosides, and gangliosides are common in mammalian tissues, especially in the brain and other nervous tissues, but are rare in most bacteria. The production of sphingolipids can be found in a number of other genera in the phylum Bacteroidetes including Flectobacillus, Prevotella, Porphyromonas, and Sphingobacterium.
Figure 16.41 Sphingolipids.

Comparison of (a) glycerol with (b) sphingosine. In sphingolipids, characteristic of Bacteroides species, sphingosine is the esterifying alcohol; a fatty acid is bonded by peptide linkage through the N atom (shown in red), and the terminal —OH group (shown in green) can be part of any of a number of compounds including phosphatidylcholine (sphingomyelin) or various sugars (cerebrosides and gangliosides).
Check Your Understanding
What is the role of Bacteroides thetaiotaomicron in the human gut?
16.14 Cytophagales, Flavobacteriales, and Sphingobacteriales
16.14 Cytophagales, Flavobacteriales, and Sphingobacteriales
*Cytophagales*
The order Cytophagales (Figure 16.40) contains almost exclusively obligate aerobes, though some species have limited fermentative capabilities. Cells are typically long, slender, gram-negative rods, often containing pointed ends, and move by gliding (Figure 16.42). Cytophagas specialize in the degradation of complex polysaccharides. They are widespread in toxic soils and freshwaters, where they probably account for much of the bacterial cellulose digestion. Cellulose decomposers can easily be isolated by placing small crumbs of soil on pieces of cellulose filter paper laid on the surface of mineral salts agar. The bacteria attach to and digest the cellulose fibers, forming spreading colonies (Figure 16.42b).
Figure 16.42 *Cytophaga* and *Sporocytophaga*.

(a) Streak of an agarolytic marine Cytophaga hydrolyzing agar in a Petri dish. (b) Colonies of Sporocytophaga growing on cellulose. Note the clearing zones (arrows) where the cellulose has been degraded. (c) Phase-contrast micrograph of cells of Cytophaga hutchinsonii grown on cellulose filter paper (cells are about 1.5 μm in diameter). (d) Phase-contrast micrograph of the rod-shaped cells and spherical microcysts of Sporocytophaga myxococcoides (cells are about 0.5 μm and microcysts about 1.5 μm in diameter). Although Sporocytophaga microcysts are only slightly more heat-tolerant than vegetative cells, they are extremely resistant to desiccation and thus help the organism survive dry periods in soil. The genera Cytophaga and Sporocytophaga form a major clade within the phylum Bacteriodetes (Figure 16.40).
Cellulose degradation by cytophagas can proceed by two different mechanisms. The typical mechanism is the free cellulase mechanism in which cells secrete extracellular enzymes called exoenzymes that degrade insoluble cellulose outside of the cell. A complex mixture of enzymes is secreted including processive endocellulases, which cleave internal β-1,4 glycosidic bonds, and processive exocellulases, which cleave terminal β-1,4 glucosidic bonds, releasing cellobiose. These exoenzymes degrade insoluble cellulose into soluble polysaccharides and disaccharides that can be readily assimilated by cells. Cytophaga hutchinsonii does not produce processive cellulases, and its degradation of cellulose likely requires physical contact of cellulose fibers with cellulase enzymes located on the outer surface of its cell wall.
The genus Cytophaga contains species that can degrade not only cellulose (Figure 16.42c) but also agar (Figure 16.42a) and chitin. In pure culture Cytophaga can be grown on agar containing embedded cellulose fibers (Figure 16.42b). The related genus Sporocytophaga is similar to Cytophaga in morphology and physiology, but the cells form resting spherical structures called microcysts (Figure 16.42d), similar to those produced by some fruiting myxobacteria (Section 15.16).
Several species of Cytophaga are fish pathogens and can cause serious problems in the cultivated fish industry. Two of the most important diseases are columnaris disease, caused by Cytophaga columnaris, and cold-water disease, caused by Cytophaga psychrophila. Both diseases preferentially affect stressed fish, such as those living in waters receiving pollutant discharges or living in high-density confinement situations such as fish hatcheries and aquaculture facilities. Infected fish show tissue destruction, frequently around the gills, probably from proteolytic activities of the Cytophaga pathogen.
*Flavobacteriales* and *Sphingobacteriales Flavobacteriales* and *Sphingobacteriales* (Figure 16.40) typically contain aerobic and facultatively aerobic chemoorganotrophs. Like most *Bacteroidetes*, these organisms are gram-negative rods, and are saccharolytic with many species motile by gliding. Species are found widely in soils and in aquatic habitats, where they typically degrade complex polysaccharides.
Flavobacteriales can be particularly abundant in marine waters including aquatic systems in polar environments. Flavobacterium species are primarily found in aquatic habitats, both freshwater and marine, as well as in foods and food-processing plants. Most species are obligate aerobes, though some species are able to reduce nitrate in an anaerobic respiration. Flavobacteria frequently produce yellow pigments and are generally saccharophilic; most can also degrade starch and proteins. Flavobacteria are rarely pathogenic; however, one species, Flavobacterium meningosepticum, has been implicated in cases of infant meningitis, and several fish pathogens are also known.
Some Flavobacteriales are psychrophilic or psychrotolerant (Section 4.12). These include, in particular, the genera Polaribacter and Psychroflexus, organisms commonly isolated from cold environments, especially permanently cold environments such as polar waters and sea ice. Many related genera are also capable of good growth below 20 °C and can thus be agents of food spoilage. None are pathogenic.
Sphingobacteriales are phenotypically similar to many Flavobacteriales. In terms of physiology, species of Sphingobacteriales are generally able to degrade a greater breadth of complex polysaccharides than are Flavobacteriales, and in this regard they resemble species of Cytophagales. The genus Flexibacter is typical of many genera of Sphingobacteriales. Species of Flexibacter differ from those of Cytophaga in that they usually require complex media for good growth and are unable to degrade cellulose. Cells of some Flexibacter species also undergo changes in cell morphology from long, gliding, threadlike filaments lacking cross-walls to short, nonmotile rods. Many flexibacteria are pigmented due to carotenoids located in their cytoplasmic membrane, or from related pigments called flexirubins, located in the cell’s outer membrane. Flexibacter species are common in soil and freshwaters where they degrade polysaccharides, and none have been identified as pathogens.
Check Your Understanding
Describe a method for isolating Cytophaga species from nature.
What characteristics are shared between the genera Cytophaga and Bacteroides, and in what ways do they differ?
IV: Chlamydiae, Planctomycetes, and Verrucomicrobia
IV: Chlamydiae, Planctomycetes, and Verrucomicrobia
IV *Chlamydiae*, *Planctomycetes*, and *Verrucomicrobia*
***C hlamydiaeandPlanctomyceteslack the key cell-division protein FtsZ and produce unusual cellular structures.Chlamydiaeinclude intracellular parasites whilePlanctomycetesandVerrucomicrobia* are widespread in soils and aquatic systems.**
The phyla Chlamydiae, Planctomycetes, and Verrucomicrobia share an ancestor and are more closely related to each other than to other bacterial phyla (Figure 16.43). These three groups contain organisms that can be found in a variety of habitats including soils, aquatic systems, and in association with eukaryotic hosts. The Chlamydiae and Planctomycetes are highly unusual among bacteria in that they lack a gene encoding the protein FtsZ, a key protein in septum formation during cell division (Section 8.3). FtsZ is essential for cell division in most bacteria and hence there is great interest in understanding how Chlamydiae and Planctomycetes divide without this protein. We first consider the chlamydias, a group of small gram-negative bacteria that cause some serious human and animal diseases.
Figure 16.43 Major orders of *Chlamydiae*, *Planctomycetes*, and *Verrucomicrobia*.

The phylogenetic tree was constructed from 16S rRNA gene sequences of representative genera of Chlamydiae, Planctomycetes, and Verrucomicrobia. Order names are shown in bold.
Key Genera: *Chlamydia, Chlamydophila, Parachlamydia*
The phylum Chlamydiae contains a single order, the Chlamydiales. The entire phylum consists of obligate intracellular parasites of eukaryotes, in particular, humans. The genomes of the chlamydias are highly reduced, being about 1 Mbp in size, and as a result they are among the most biochemically limited of all known Bacteria. They are unable to synthesize many of the vitamins and growth factors needed to grow and must acquire these molecules from the cytoplasm of infected host cells.
Besides their lack of the key protein FtsZ (Section 8.3), many chlamydial species lack a cell wall. These organisms apparently minimize their use of peptidoglycan as a means of remaining undetected by the host immune system. Despite the complete absence of a cell wall in many species of the Chamydiae these organisms remain susceptible to beta-lactam antibiotics such as penicillin, a drug that prevents peptidoglycan synthesis. The answer to this conundrum is the fact that although they lack a cell wall, chlamydias still require the formation of a ringlike peptidoglycan structure to facilitate septum formation during cell division. In the absence of this structure, cell division is compromised.
Though the chlamydia that are human pathogens have been characterized in most detail, the phylum contains diverse species that interact with a wide variety of eukaryotic hosts including diverse animals and even many amoebae. Species are typically very small cocci, approximately 0.5 μm in diameter, and display a distinctive developmental cycle.
Life Cycle of *Chlamydiae*
Species of Chlamydiae demonstrate a unique chlamydial life cycle (Figure 16.44). Two types of cells are seen in the life cycle: (1) a small, dense cell, called an elementary body, which is relatively resistant to drying and is the means of dispersal, and (2) a larger, less dense cell, called a reticulate body, which divides by binary fission and is the vegetative form.
Figure 16.44 The infection cycle of a chlamydia.

(a) Schematic diagram of the cycle: The entire cycle takes about 48 h. (b) Human chlamydial infection. Elementary bodies (∼0.3 μm in diameter) are the infectious form and reticulate bodies (∼1 μm in diameter) are the multiplying form. An infected fallopian tube cell is bursting, releasing mature elementary bodies.
Elementary bodies are nonmultiplying cells specialized for infectious transmission. By contrast, reticulate bodies are noninfectious forms that function only to multiply inside host cells to form a large inoculum for transmission. Unlike the rickettsias, the chlamydias are not transmitted by arthropods but are primarily airborne invaders of the respiratory system—hence the significance of resistance to drying of the elementary bodies. A dividing reticulate body can be seen in Figure 16.45. After a number of cell divisions, these vegetative cells are converted into elementary bodies that are released when the host cell disintegrates (Figure 16.44b) and can then infect other nearby host cells. Generation times of 2–3 h have been measured for reticulate bodies, considerably faster than times found for the rickettsias (Section 16.1).
Figure 16.45 *Chlamydiae*.

Thin-section electron micrograph of a dividing reticulate body of Chlamydophila psittaci within a mouse tissue-culture cell. A single chlamydial cell is about 1 μm in diameter.
Mastering Microbiology
Art Activity: Figure 16.44a The infection cycle of a chlamydia
The best-studied human pathogens are found in the genera Chlamydia and Chlamydophila. Several species are recognized within these genera: Chlamydophila psittaci, the causative agent of the disease psittacosis; Chlamydia trachomatis, the causative agent of trachoma and a variety of other human diseases; and Chlamydophila pneumoniae, the cause of some respiratory syndromes. Psittacosis is an epidemic disease of birds that is occasionally transmitted to humans and causes pneumonia-like symptoms. Trachoma, a debilitating disease of the eye characterized by vascularization and scarring of the cornea, is the leading cause of blindness in humans. Other strains of C. trachomatis infect the genitourinary tract, and chlamydial infections are currently one of the leading sexually transmitted diseases ( Section 31.13).
In sum, the chlamydias appear to have evolved an efficient and effective survival strategy including parasitizing the resources of the host and producing resistant cell forms specialized for transmission.
Check Your Understanding
How are Chlamydia and Mycoplasma (Section 16.9) similar? How are they different?
What is the difference between an elementary body and a reticulate body?
Key Genera: *Planctomyces, Blastopirellula, Gemmata, Brocadia*
The phylum Planctomycetes contains a diverse set of morphologically unique bacteria found primarily in two orders, Planctomycetales and Brocadiales (Figure 16.43). Planctomycetes are gram-negative bacteria that often have stalks or appendages, and many species form rosettes (clusters of cell typically attached at their poles). Planctomycetes are found widely in soils and aquatic environments and they are rarely pathogenic. Despite being observed widely in the environment, their ecology remains poorly defined. Planctomycetes are remarkable for having a set of unusual cellular features.
Unlike most Bacteria, Planctomycetes lack the protein FtsZ (see also Chlamydia, Section 16.15), and they divide by budding, much like yeast. Moreover, Planctomycetes were long thought to lack peptidoglycan, and transmission electron micrographs (TEM) suggested the presence of a membrane-enclosed nucleus in some species (Figure 16.46). This unique mix of prokaryotic and eukaryotic features has for years given the impression that Planctomycetes are a missing link in cellular evolution. However, the mysteries of the missing peptidoglycan and the nature of the “nucleus”—like many other microbial mysteries—were ultimately solved by advances in microbiological techniques.
Figure 16.46 Membrane invaginations in *Gemmata*.

Thin-section electron micrograph of a cell of Gemmata obscuriglobus showing the nucleoid surrounded by an invagination of the cell membrane (see Figure 16.47). The deep invagination of the cell membrane, when viewed in a two-dimensional section, gives the appearance of a nuclear membrane. The cell is about 1.5 μm in diameter.
Compartmentalization in *Planctomycetes*
Cryo-electron tomography (CET) is a microscopic method that generates detailed three-dimensional images with nanometer-scale resolution (Section 1.10). CET scans of Planctomycetes revealed that what appeared to be a nuclear membrane in two-dimensional transmission electron micrographs was actually a system of deep invaginations of the cytoplasmic membrane (Figure 16.47). The cytoplasmic membrane of cells of Planctopirus is heavily invaginated, and when viewed in three dimensions, these invaginations can be seen to wrap around the DNA-containing nucleoid (Figure 16.47). Two-dimensional slices through this three-dimensional system of membranes gave the false appearance of a nuclear membrane (for example, see Figure 16.46). Furthermore, these images clearly reveal that Planctomycetes have a typical gram-negative cell envelope containing peptidoglycan.
Figure 16.47 Cryo-electron tomographic reconstruction of *Planctopirus*.

Planctomycetes have a complex cell structure consisting of a series of deep and complex invaginations of the cellular membrane. Until recently, these unusual membrane structures were interpreted as evidence that Planctomycetes could have a membrane-bound nucleus (see Figure 16.46). The image shows the internal structure of a single cell and the outer sections of several nearby cells. The structures are color coded: outer membrane (green), a peptidoglycan-containing cell wall (orange), cytoplasmic membrane (blue), DNA-containing nucleoids (yellow), ribosomes (white) present in the cytoplasm, and crateriform structures (magenta) on the outer membrane. The presence of two nucleoids suggest the central cell is preparing to divide.
While we now know that cells of Planctomycetes contain both peptidoglycan and a gram-negative cell envelope, an understanding of the remarkable membrane invaginations that appear in CET scans of cells of Planctomycetes remains elusive. In addition, these CET scans reveal unusual crateriform structures studding the surface of their outer membranes. It is possible that these crateriform structures mediate the transport of large molecules into the cell where they are then metabolized in the large periplasmic compartments formed by the membrane invaginations. However, the purpose of these cellular features remains unknown.
Another interesting cellular structure found in the Planctomycetes is present in the Brocadiales that carry out the anammox process (Section 14.10). Anammox bacteria such as Brocadia anammoxidans contain a true organelle called the anammoxosome (Figure 14.26). These bacteria catalyze the anaerobic oxidation of ammonia (NH3) within the anammoxosome structure. The anammoxosome membrane is composed of unique lipids that form a tight seal, protecting cytoplasmic components from toxic intermediates produced during the anaerobic oxidation of ammonia (Section 14.10 and Figure 14.26c). The evolutionary origins of this organelle and many aspects of its function remain unknown.
Figure 16.48 *Planctomyces maris*.

Metal-shadowed transmission electron micrograph. A single cell is about 1–1.5 μm long. Note the fibrillar nature of the stalk. Pili are also abundant. Note also the flagella (curly appendages) on each cell and the bud that is developing from the nonstalked pole of one cell.
Like Caulobacter (Figures 8.6, 8.20, and 15.54), Planctomyces is a budding bacterium that displays a life cycle. Its motile swarmer cells attach to a surface, grow a stalk from the attachment point, and generate a new cell from the opposite pole by budding. This daughter cell produces a flagellum, breaks away from the attached mother cell, and begins the cycle anew. Physiologically, Planctomyces species are facultatively aerobic chemoorganotrophs, growing either by fermentation or respiration of sugars.
Planctomyces are found primarily in soil and aquatic habitats, both freshwater and marine, and the genus Isosphaera is a filamentous, gliding hot spring bacterium. The isolation of Planctomyces and relatives, like that of Caulobacter, requires dilute media.
Check Your Understanding
How does the stalk of Planctomyces differ from the stalk of Caulobacter?
What is unusual about the bacterium Planctopirus?
Key Genera: *Verrucomicrobium, Prosthecobacter*
The phylum Verrucomicrobia contains at least four orders with characterized species, but most are found within the order Verrucomicrobiales (Figure 16.43). Species of Verrucomicrobia are aerobic or facultatively aerobic bacteria capable of fermenting sugars. An exception is the genus Methylacidiphilum, which contains aerobic methanotrophs (Section 15.15). In addition, some Verrucomicrobia form symbiotic associations with protists. Verrucomicrobia are widespread in nature, inhabiting freshwater and marine environments as well as forest and agricultural soils. The Verrucomicrobia can have membrane-bound intracellular structures similar to those found in the Planctomycetes. The Verrucomicrobia typically form cytoplasmic appendages called prosthecae (Section 15.18). Verrucomicrobia share with other prosthecate bacteria the presence of peptidoglycan in their cell walls and in this way are clearly distinct from Planctomycetes.
The genera Verrucomicrobium and Prosthecobacter produce two to several prosthecae per cell (Figure 16.49). Unlike cells of Caulobacter (Figures 8.6, 8.20, and 15.54), which contain a single prostheca and produce flagellated and nonprosthecate swarmer cells, Verrucomicrobium and Prosthecobacter divide symmetrically, and both mother and daughter cells contain prosthecae at the time of cell division. The genus name Verrucomicrobium derives from Greek roots meaning “warty,” which is an appropriate description of cells of Verrucomicrobium spinosum with their multiple projecting prosthecae (Figure 16.49).
Figure 16.49 *Verrucomicrobium spinosum*.

Negatively stained transmission electron micrograph. Note the wartlike prosthecae. A cell is about 1 μm in diameter.
Mastering Microbiology
Species of the genus Prosthecobacter contain two genes that show significant homology to the genes that encode tubulin in eukaryotic cells. Tubulin is the key protein that makes up the cytoskeleton of eukaryotic cells (Section 2.15). Although the important cell-division protein FtsZ (Section 8.3) is also a tubulin homolog, the Prosthecobacter proteins are structurally more similar to eukaryotic tubulin than is FtsZ. The role of the tubulin proteins in Prosthecobacter is unknown since a eukaryotic-like cytoskeleton has not been observed in these organisms.
Check Your Understanding
Describe two ways that Verrucomicrobia differ from Planctomycetes.
V Hyperthermophilic *Bacteria*
***Aquificales* are common in geothermally heated aquatic systems. Hyperthermophilic phyla branch near the root of the bacterial tree, supporting the idea that Earth’s first bacterial cells were thermophilic.**
Three phyla of hyperthermophilic bacteria cluster deep in the phylogenetic tree of Bacteria, near the root (Figure 16.1). Each group consists of one or two major genera, and a key physiological feature of most species is hyperthermophily—optimal growth at temperatures above 80 °C (Section 4.13). We begin with Thermotoga and Thermodesulfobacterium, each representative of its own lineage.
Key Genera: *Thermotoga, Thermodesulfobacterium*
The phylum Thermotogae includes about 12 genera containing cultivated organisms. These species are all chemoorganotrophic anaerobes and many are thermophiles and hyperthermophiles, though some mesophiles are known. Species of the representative genus Thermotoga are rod-shaped hyperthermophiles that form a sheathlike envelope (called a toga; thus the genus name) (**Figure 16.50*a***), stain gram-negatively, and are nonsporulating. Thermotoga species are fermentative anaerobes, catabolizing sugars or starch and producing lactate, acetate, CO2, and H2 as fermentation products. The organisms can also grow by anaerobic respiration using H2 as an electron donor and ferric iron as an electron acceptor. Species of Thermotoga have been isolated from terrestrial hot springs as well as marine hydrothermal vents.
Figure 16.50 Hyperthermophilic *Bacteria*.

Electron micrographs of two hyperthermophiles: (a) Thermotoga maritima—temperature optimum, 80 °C. Note the outer covering, the toga. (b) Aquifex pyrophilus—temperature optimum, 85 °C. Cells of Thermotoga measure 0.6×3.5 μm; cells of Aquifex measure 0.5×2.5 μm.
Despite being bacterial, the genome of Thermotoga contains many genes that show strong homology to genes from hyperthermophilic Archaea. In fact, over 20% of the genes of Thermotoga probably originated from Archaea by horizontal gene transfers (Section 13.9 and Chapter 9). Although a few archaeal-like genes have been identified in the genomes of other Bacteria and vice versa, only in Thermotoga has such large-scale horizontal transfer of genes between domains been detected thus far.
Thermodesulfobacterium (Figure 16.51) is a thermophilic sulfate-reducing bacterium, positioned on the phylogenetic tree in a separate phylum after Thermotoga and Aquifex (Figure 16.1a). Thermodesulfobacterium is a strict anaerobe that uses compounds such as lactate, pyruvate, and ethanol (but not acetate) as electron donors, as do sulfate-reducing bacteria such as Desulfovibrio (Section 15.11), reducing SO4 2− to H2S.
Figure 16.51 *Thermodesulfobacterium*.

(a) Phase-contrast micrograph of cells of Thermodesulfobacterium thermophilum. (b) Structure of one of the lipids of Thermodesulfobacterium mobile. Note that although the two hydrophobic side chains are ether-linked, they are not phytanyl units, as in Archaea. The designation “R” is for a hydrophilic residue, such as a phosphate group.
An unusual biochemical feature of Thermodesulfobacterium is the production of ether-linked lipids. Recall that such lipids are a hallmark of the Archaea and that a polyisoprenoid C20 hydrocarbon (phytanyl) replaces fatty acids as the side chains in archaeal lipids (Section 2.1). However, the ether-linked lipids in Thermodesulfobacterium are unusual because the glycerol side chains are not phytanyl groups, as they are in Archaea, but instead are composed of unique C17 hydrocarbons along with some fatty acids (Figure 16.51b). Thus we see in Thermodesulfobacterium both a deep phylogenetic lineage (Figure 16.1) and a lipid profile that combines features of both the Archaea and the Bacteria. However, a few other Bacteria have also been found to contain ether-linked lipids, and thus these lipids may be more common among Bacteria than previously thought.
Check Your Understanding
What is unique about the genome of Thermotoga and the lipids of Thermodesulfobacterium?
Key Genera: *Aquifex, Thermocrinis*
The genus Aquifex (Figure 16.50b) is an obligately chemolithotrophic and autotrophic hyperthermophile and is the most thermophilic of all known Bacteria. Various Aquifex species utilize H2, sulfur (S0), or thiosulfate (S2O3 2−) as electron donors and O2 or nitrate (NO3 −) as electron acceptors, and grow at temperatures up to 95 °C. Aquifex can tolerate only very low O2 concentrations (microaerophilic), and is unable to oxidize any tested organic compound. Hydrogenobacter, a relative of Aquifex, shows most of the same properties as Aquifex, but is an obligate aerobe.
*Aquifex* and Autotrophy
Autotrophy in Aquifex occurs by way of the reverse citric acid cycle, a series of reactions previously detected only in green sulfur bacteria (Sections 14.2 and 15.6) within the domain Bacteria. The genome sequence of Aquifex aeolicus reveals that an entirely chemolithotrophic and autotrophic lifestyle is encoded by a genome of only 1.55 Mbp (one-third the size of the Escherichia coli genome). The discovery that so many hyperthermophilic species of Archaea and Bacteria, like Aquifex, are H2 chemolithotrophs, coupled with the finding that they branch as very early lineages on their respective phylogenetic trees (Figure 16.1a), suggests that H2 was a key electron donor for energy metabolism in primitive organisms that appeared on early Earth and also suggests that the earliest cells may have been thermophiles (Section 13.1 and Section 17.14).
Figure 16.52 *Thermocrinis*.

(a) Octopus Spring, Yellowstone National Park (USA). The source water of this alkaline and siliceous hot spring is 92 °C. (b) Cells of Thermocrinis ruber growing as filamentous streamers (arrow) attached to siliceous sinter in the outflow (85 °C) of Octopus Spring (Figure 17.19b). (c) Scanning electron micrograph of rod-shaped cells of T. ruber grown on a silicon-coated cover glass. A single cell of T. ruber is about 0.4 μm in diameter and from 1 to 3 μm long.
T. ruber is of historical significance in microbiology because it was one of the organisms discovered in the 1960s by Thomas Brock, a pioneer in the field of thermal microbiology and senior author of the first seven editions of this text. The discovery by Brock that the pink streamers (Figure 16.52b) contained protein and nucleic acids clearly indicated that they were living organisms and not just mineral debris. Moreover, the presence of streamers in 80–90 °C hot spring outflow waters but not those of lower temperatures supported Brock’s hypothesis that these organisms actually required heat for growth and were therefore likely to be present in even boiling or superheated waters. Both of these conclusions were subsequently supported by the discovery by Brock, Karl Stetter of Germany, and many other microbiologists of dozens of genera of hyperthermophilic Bacteria and Archaea inhabiting hot springs, hydrothermal vents, and other thermal environments.More coverage of hyperthermophiles can be found in Section 4.13 and Chapter 17.
Check Your Understanding
Of what evolutionary significance is the fact that organisms in the Aquifex lineage are both hyperthermophilic and H2 chemolithotrophs?
VI Other *Bacteria*
***Bacteriasuch asAcidobacteriaandFusobacteriaare difficult to cultivate but are abundant and important components of various natural habitats and the human microbiome, respectively.DeinococcusandThermus* can survive extremes of radiation and temperature, respectively.**
Thus far in this chapter we have focused on phyla that have many described species (Figure 16.1). Beyond these mainstream bacterial phyla are many others that have but one or at most a handful of characterized species (Figure 16.1b). In addition, many more phyla are known only from community sampling of 16S rRNA genes from nature (Sections 19.6 and 19.8). We cannot cover them all. So in this final part of the chapter we consider two more phyla that have been well studied and then summarize some other phyla that are emerging into the mainstream of microbial diversity.
Key Genera: *Deinococcus, Thermus*
The deinococci group contains only a few characterized genera in two orders, the Deinococcales and the Thermales. Members of this phylum are typically aerobic chemoorganotrophs that metabolize sugars, amino and organic acids, or various complex mixtures. Though deinococci stain gram-positively, they have a gram-negative cell wall structure (Figure 16.53) made up of several layers, including an outer membrane, which is characteristic of gram-negative bacteria (Section 2.3). However, unlike the outer membrane of bacteria such as Escherichia coli, the outer membrane of deinococci lack lipid A. Deinococci also contain an unusual form of peptidoglycan in which ornithine replaces diaminopimelic acid in the N-acetylmuramic acid cross-links (Section 2.3).
Figure 16.53 The radiation-resistant coccus *Deinococcus radiodurans*.

An individual cell is about 2.5 μm in diameter. (a) Transmission electron micrograph of D. radiodurans. Note the outer membrane layer. (b) High-magnification micrograph of wall layer. (c) Transmission electron micrograph of cells of D. radiodurans colored to show the toroidal morphology of the nucleoid (green).
Species of Thermales are typically thermophiles or hyperthermophiles and the type genus is Thermus. Thermus aquaticus, discovered in a Yellowstone National Park hot spring in the mid-1960s by Thomas Brock (Section 16.19), has been a model organism for studying life at high temperatures. T. aquaticus has subsequently been isolated from many thermal systems including private and commercial hot water heaters and is the source of Taq DNA polymerase. Because it is so heat-stable, Taq polymerase allowed the polymerase chain reaction (PCR) technique for amplifying DNA to be fully automated (Section 12.1), an advance that revolutionized all of biology.
Radiation Resistance of *Deinococcus radiodurans*
Species of Deinococcales have the unusual property of being extremely radiation resistant, and Deinococcus radiodurans is the best-studied species in this regard. Most deinococci are red or pink due to carotenoids, and many are highly resistant to both radiation and desiccation. Resistance to ultraviolet (UV) radiation can be used to advantage in isolating deinococci. These remarkable organisms can be selectively isolated from soil, ground meat, dust, and filtered air following exposure of the sample to intense UV (or even gamma) radiation and plating on a rich medium containing tryptone and yeast extract. For example, D. radiodurans cells survive exposure to 15,000 grays (Gy) of ionizing radiation (1 Gy=100 rad). This is sufficient to shatter the organism’s chromosome into hundreds of fragments (by contrast, some endospores are killed by lower levels, and a human can be killed by exposure to less than 10 Gy) (Section 4.18).
In addition to impressive radiation resistance, D. radiodurans is resistant to the effects of many mutagenic agents. The only chemical mutagens that seem to work on D. radiodurans are agents such as nitrosoguanidine, which induces deletions in DNA. Deletions are apparently not repaired as efficiently as point mutations in this organism, and mutants of D. radiodurans can be isolated in this way.
DNA Repair in *Deinococcus radiodurans*
Studies of D. radiodurans have shown that it is highly efficient in repairing damaged DNA. Several different DNA repair enzymes exist in D. radiodurans. In addition to the DNA repair enzyme RecA (Sections 9.4 and 9.5), several RecA-independent DNA systems exist in D. radiodurans that can repair breaks in single- or double-stranded DNA, and excise and repair misincorporated bases. In fact, repair processes are so effective that the chromosome can even be reassembled from a fragmented state.
It is also thought that the unique arrangement of DNA in D. radiodurans cells plays a role in radiation resistance. Cells of D. radiodurans always exist as pairs or tetrads (Figure 16.53a). Instead of scattering DNA within the cell as in a typical nucleoid, DNA in D. radiodurans is ordered into a toroidal (coiled, or stack of rings) structure (Figure 16.53c). Repair is then facilitated by the fusion of nucleoids from adjacent compartments, because their toroidal structure provides a platform for homologous recombination. From this extensive recombination, a single repaired chromosome emerges, and the cell containing this chromosome can then grow and divide.
Check Your Understanding
Describe a commercial application of Thermus aquaticus.
Describe an unusual biological feature of Deinococcus radiodurans.
16.21 *Acidobacteria* and *Nitrospirae*
Surveys of environmental DNA (Sections 19.6 and 19.8) reveal Acidobacteria and Nitrospirae to be abundant and widespread in the environment (Figure 16.1b). Despite their abundance, strains of these phyla have proven difficult to cultivate and much of our knowledge about the Acidobacteria and Nitrospirae comes from metagenomic surveys that detect their genes in natural samples (Section 10.7). We begin with the Acidobacteria, which are notable for their diversity and abundance in soils (Section 20.7 and Figure 20.14).
Key Genus: *Acidobacterium Acidobacteria* are widespread in the environment as revealed by analyses of 16S rRNA genes (Figure 16.1*b*). *Acidobacteria* are common in soils, and especially in acid soils (pH<6.0), where they often comprise a majority of soil microbial communities. *Acidobacteria* also inhabit freshwater, hot spring microbial mats, wastewater treatment reactors, and sewage sludge. Their abundance, widespread distribution, and phylogenetic diversity suggest they play important ecological roles, especially in soil. Unfortunately, while *Acidobacteria* are widespread in the environment, few have been cultivated (Figure 16.1*b*) and only a handful of genera have been described.
Most described species of Acidobacteria are gram-negative, acid-tolerant, aerobic chemoorganotrophs. For example, Acidobacterium capsulatum is an acidophile isolated from acid mine drainage. An obligately aerobic encapsulated bacterium, it grows using various sugars and organic acids. Acidisarcina polymorpha, which grows in cuboidal packets of 6–8 cells (Figure 16.54), was isolated from an acidic boreal forest soil and is also acid-tolerant. This bacterium is aerobic and able to degrade polymers such as cellulose and chitin, which are common in soils. Pyrinomonas methylaliphatogenes is a thermophile isolated from thermally heated soil. This species is also an acidophilic aerobic chemoorganotroph. P. methylaliphatogenes is notable, however, because it can generate energy, but not grow, using atmospheric H2, typically present in oxic environments in only trace amounts. It is unclear whether this ability is widespread among Acidobacteria, but if so, the ability to survive prolonged starvation might explain why Acidobacteria are so common in soils.
Figure 16.54 Cells of *Acidisarcina polymorpha*.

This species of Acidobacteria was isolated from acidic Russian soils and degrades both cellulose and chitin, polymers that are common in soil. Cells typically occur in cuboid packets containing to 6–8 cells encapsulated in a shared polysaccharide (see Sarcina, Figure 16.21). Both phase-contrast (a) and TEM (b) images are shown. Cells have an irregular shape and are 1.4–4.4 μm long.
Acidophilic, aerobic chemoorganotrophs are common among cultivated strains of Acidobacteria, but this phylum contains tremendous phylogenetic diversity spanning at least 26 major subgroups. Not all of these subgroups appear to contain acidophilic organisms and not all are aerobic chemoorganotrophs. For example, Geothrix fermentans is a strict anaerobe capable of dissimilative iron reduction (Section 15.13) and can also ferment citrate to acetate plus succinate. Holophaga foetida is a strictly anaerobic acetogen (Section 14.14) that grows by degrading methylated aromatic compounds to acetate. Finally, at least one genus, Chloracidobacterium, is phototrophic (Section 15.8).
Key Genus: *Nitrospira*
The phylum Nitrospirae is named for the genus Nitrospira, a chemolithotroph that oxidizes nitrite to nitrate and grows autotrophically, as do species of the proteobacterium Nitrobacter (Section 15.10). Nitrospira inhabits many of the same environments as Nitrobacter. However, environmental surveys have shown that Nitrospira is much more abundant than Nitrobacter in nature, and thus most of the nitrite oxidized in nitrogen-rich environments such as wastewater treatment plants and ammonia-rich soils is probably due to Nitrospira. Some species of Nitrospira found widely in soils have also been observed to contain the complete pathway for nitrification, oxidizing ammonia all the way to nitrate (Sections 14.9, 15.10, and Section 21.3). Other key Nitrospirae include Leptospirillum, an aerobic, acidophilic, iron-oxidizing chemolithotroph (Section 15.14) common in acid mine drainage associated with the mining of coal and iron (Section 22.1).
Check Your Understanding
In what kind of habitat would you expect to find Acidobacteria in high abundance? Make a hypothesis as to why Acidobacteria are favored in such habitats.
What different reactions can Nitrospira species perform during the process of nitrification?
16.22 Other Notable Phyla of *Bacteria*
The basic properties of several other phyla of Bacteria are discussed briefly below. Although most of these phyla have few cultured representatives (Figure 16.1b), many may well be of considerable importance. If so, future research on their culture and ecological activities will supply the necessary proof. Until then, we paint a picture of these phyla with a broad brush to summarize their major characteristics in a general way.
Fusobacteria
Fusobacteria are gram-negative nonsporulating rods found in sediments and also the gastrointestinal systems and oral cavities of animals. Fusobacteria are obligate anaerobes that ferment carbohydrates, peptides, and amino acids. Species of the genus Fusobacterium are a common component of the human microbiome (Chapter 24). Fusobacteria have adhesions, molecules that allow them to form strong attachments to other cells, both bacterial and animal. These adhesions allow Fusobacteria to play an important structural role in the formation of biofilms called dental plaque in the oral cavity (Figure 16.55). Fusobacteria are difficult to cultivate and so their role in the human microbiome was not fully appreciated until the development of DNA-based techniques for studying microbial communities (Section 19.8).
Figure 16.55 Cells of *Fusobacterium* in human dental plaque.

The photo is a light micrograph showing a section of human dental plaque stained by fluorescence in situ hybridization (FISH, Section 19.5). While streptococci (Section 16.6), stained green here, are common in plaque and are the primary cause of dental caries (cavities, Section 24.3), plaque is a complex biofilm composed of many different species (each in different colors, see Figures 24.10 and 25.8 and the Chapter 19 MicrobiologyNow). Fusobacterium, stained yellow here, are filamentous bacteria with adhesins that allow them to attach to other cells. This trait allows them to colonize cellular surfaces and play a role in structurally supporting these complex biofilms. Fusobacteria grow within the biofilm along with other anaerobic filamentous organisms providing an anchor for lactic acid bacteria that grow closer to the biofilm surface.
While Fusobacteria are common in the microbiome of healthy individuals (Chapter 24), they are also implicated in several human diseases and syndromes. In particular, Fusobacteria increase in abundance in individuals suffering from a range of oral diseases. Inflammation of gum tissue and tears in blood vessels allow Fusobacteria and other oral bacteria access to the bloodstream. Such bacteria can be found in arterial plaques and may even play a role in heart disease. In addition, Fusobacteria in the bloodstream have also been shown to cause adverse pregnancy outcomes. Finally, Fusobacteria in the gut have been shown to attach, colonize, and enhance the growth of colorectal cancer cells. In mice with such tumors, treatment with the antibiotic metronidazole both reduced the abundance of Fusobacteria in tumors and reduced tumor growth. It would seem that we still have a lot to learn about the Fusobacteria.
*Fibrobacteres* and *Synergistetes Fibrobacteres* and *Synergistetes* contain relatively few characterized species (Figure 16.1*b*), but those that have been cultured employ fermentative metabolisms similar to *Fusobacteria*. Species in these groups are often associated with the gastrointestinal tracts of animals.
While 16S ribosomal RNA genes from Fibrobacteres can be recovered from a wide range of habitats, the only characterized species have come from either the rumen or gastrointestinal tracts of animals. The genus Fibrobacter contains gram-negative fermentative strict anaerobes. However, unlike most Fusobacteria and Synergistetes, species of Fibrobacter are unable to ferment proteins or amino acids and specialize instead in the fermentation of carbohydrates, including cellulose. In the rumen, cellulose is the major source of energy, and in such environments it supports not only cellulolytic bacteria such as Fibrobacter but many noncellulolytic anaerobes that use glucose released during cellulose degradation.
Synergistetes are gram-negative nonsporulating rods found in association with animals and in anoxic environments in terrestrial and marine systems. Described species are typically obligate anaerobes that degrade proteins and are capable of fermenting amino acids. In animals they are most often found in the gastrointestinal tract; for example, Synergistes jonesii inhabits the rumen (Section 23.15). In humans, species of Synergistetes have been associated with certain soft tissue wounds and abscesses, dental plaque, and periodontal conditions.
*Deferribacteres* and *Chrysiogenetes*
The phyla Deferribacteres and Chrysiogenetes (Figure 16.1) contain anaerobic chemoorganotrophs that display considerable metabolic diversity with respect to the electron acceptors used in anaerobic respirations (Chapter 14 and Figure 14.1). Most, though not all, species are able to grow through anaerobic respiration of nitrate to nitrite or ammonium. The Deferribacteres group is named for the genus Deferribacter, a thermophilic dissimilative ferric iron-reducer (Sections 14.13 and 15.13) that can also reduce nitrate and metal oxides. Geovibrio is a related genus that can also grow using elemental sulfur as an electron acceptor (Section 15.11). The bacterium Chrysiogenes arsenatis and its relatives are notable for the ability to couple the oxidation of acetate and a few other organic compounds to the reduction of arsenate as a terminal electron acceptor, reducing it to arsenite. In addition to arsenate, many species of Chrysiogenetes can reduce selenate, nitrite, nitrate, thiosulfate, and elemental sulfur in anaerobic respirations (Section 14.13).
Check Your Understanding
How do Nitrospira (Section 16.21) and Deferribacter differ in terms of lifestyle and metabolism?
What metabolic characteristics are shared by most Fusobacteria, Fibrobacteres, and Synergistetes, and what disease in humans has been correlated with the presence of Synergistetes and Fusobacteria?
Chapter Review
Go to Mastering Microbiology for videos, animations, practice tests, and more.
I *Proteobacteria*
16.1 The Alphaproteobacteria are the second largest class of Proteobacteria and metabolically diverse. Key genera are Rhizobium, Rickettsia, Rhodobacter, and Caulobacter.
**Q Which genera of Alphaproteobacteria are known to form nitrogen-fixing nodules in plants?**
16.2 The Betaproteobacteria are the third largest class of Proteobacteria and metabolically diverse. Key genera are Burkholderia, Rhodocyclus, Neisseria, and Nitrosomonas.
**Q What are the six major orders within the Betaproteobacteria?**
16.3 The Gammaproteobacteria are the largest and most diverse class of Proteobacteria and contain many human pathogens. The Enterobacteriales, or enteric bacteria, are the most heavily studied of all bacteria. Key genera are Escherichia and Salmonella.
Q What is the catalase test? What catalase reaction would you expect from an obligate aerobe? What reaction would you expect from an obligate anaerobe?
16.4 The Pseudomonadales and Vibrionales are among the most common Gammaproteobacteria. Key genera are Pseudomonas and Vibrio.
**Q What morphological and physiological features distinguish Escherichia from Vibrio?**
16.5 The Deltaproteobacteria and Epsilonproteobacteria are smaller and less metabolically diverse classes of Proteobacteria. Key genera of Deltaproteobacteria are Myxococcus, Desulfovibrio, and Geobacter. Key genera of Epsilonproteobacteria are Campylobacter and Helicobacter.
**Q What types of metabolism are typically associated with members of the Deltaproteobacteria and Epsilonproteobacteria, respectively?**
II *Firmicutes*, *Tenericutes*, and *Actinobacteria*
16.6 Lactic acid bacteria such as Lactobacillus and Streptococcus produce lactate as the primary end product of fermentation, and they have many roles in food production and preservation. The Firmicutes are one of the two main phyla of gram-positive bacteria.
Q What is the difference between homofermentative and heterofermentative lactic acid bacteria, and what genera are commonly associated with each of these types?
16.7 Many genera of Firmicutes in the orders Bacillales and Clostridiales, including Staphylococcus, Listeria, and Sarcina, are unable to form endospores.
**Q What characteristics of Listeria make it a frequent cause of foodborne illness?**
16.8 Production of endospores is a hallmark of the key genera Bacillus and Clostridium and is only found in the phylum Firmicutes.
Q What is a good strategy for isolating spore-forming bacteria from an environmental sample?
16.9 The phylum Tenericutes contains the mycoplasmas, organisms that lack cell walls and have very small genomes. Many species are pathogenic for humans, other animals, and plants. The key genus is Mycoplasma.
**Q Provide an explanation for why the absence of a cell wall might be favorable to Mycoplasma.**
16.10 Actinobacteria are the second major phylum of gram-positive bacteria. Corynebacterium and Arthrobacter are common gram-positive soil bacteria. Propionibacterium ferments lactate to propionate and is the key agent responsible for the unique flavor and texture of Swiss cheese.
**Q In what sort of environment would you expect to find large numbers of Actinobacteria?**
16.11 Species of Actinobacteria in the genus Mycobacterium are mainly harmless soil saprophytes, but Mycobacterium tuberculosis causes the disease tuberculosis.
**Q How does the cell wall of Mycobacterium influence its reaction to the Gram stain and the acid-fast stain?**
16.12 The streptomycetes are a large group of filamentous, gram-positive bacteria that form spores at the end of aerial filaments and are found in the phylum Actinobacteria. Many clinically useful antibiotics such as tetracycline and neomycin have come from Streptomyces species.
Q How are the spores of streptomycetes different from endospores?
III *Bacteroidetes*
16.13 The phylum Bacteroidetes includes gram-negative rods that do not form spores, many of which have gliding motility. Most species in the order Bacteroidales are obligate anaerobes that ferment carbohydrates in anoxic environments. The genus Bacteroides contains species that are common in the gastrointestinal tract of animals.
**Q What species of Bacteroidetes is most abundant in the human gastrointestinal tract, and what role does this organism play in the human gut?**
16.14 The Cytophagales and Flavobacteriales are orders in the Bacteroidetes that include aerobic bacteria able to degrade complex polysaccharides such as cellulose. These bacteria are important in organic matter decomposition.
Q What is an exoenzyme and why are these types of organisms important in the degradation of cellulose?
IV *Chlamydiae, Planctomycetes*, and *Verrucomicrobia*
16.15 The phylum Chlamydiae includes small obligate intracellular parasites that lack FtsZ and are adept at invading eukaryotic cells. Many species cause various diseases in humans and other animals.
**Q Describe the infection cycle of Chlamydia.**
16.16 The Planctomycetes are a group of stalked, budding bacteria that lack FtsZ and have extensive invaginations of the cytoplasmic membrane.
Q What is an anammoxosome and where would you expect to find one?
16.17 Species of Verrucomicrobia are distinguished by the multiple prosthecae on their cells and their unique phylogeny.
**Q Verruca is a word of Latin origin that means “wart.” How do you think the Verrucomicrobia got their name?**
V Hyperthermophilic *Bacteria*
16.18 Thermotogae and Thermodesulfobacteria form two deeply branching phyla within the Bacteria. These hyperthermophilic bacteria have proven that extensive horizontal gene transfer has occurred from Archaea to Bacteria (Thermotoga) and that ether-linked lipids are not limited to the Archaea (Thermodesulfobacterium).
**Q What major physiological property unites species of Thermotoga, Aquifex, and Thermocrinis?**
16.19 The Aquificae phylum contains a group of hyperthermophilic, H2-oxidizing bacteria that form the earliest branch on the tree of the domain Bacteria.
**Q In what environment might you observe Thermocrinis ruber, and what role did this organism play in the discovery of hyperthermophiles?**
VI Other *Bacteria*
16.20 Deinococcus and Thermus are the major genera in a distinct phylum of Bacteria. Thermus is the source of the key enzyme in automated PCR, whereas Deinococcus is the most radiation-resistant bacterium known, exceeding even many endospores in this regard.
**Q What are some of the remarkable properties that allow Deinococcus to survive exposure to massive doses of radiation?**
16.21 Acidobacteria are widespread in many environments, especially acidic soils. While aerobic, acidophilic, chemoorganotrophs are common, the phylum is metabolically and phylogenetically diverse. The genus Nitrospira includes ammonia and nitrite-oxidizing bacteria.
**Q List four ways in which different species of Acidobacteria have been shown to generate energy.**
16.22 Species of Deferribacteres and Chrysiogenetes specialize in various forms of anaerobic respiration. Species of Synergistetes, Fusobacteria, and Fibrobacteres are fermentative anaerobes that inhabit the gastrointestinal tract and other anoxic sites in animals.
**Q How is it that Fusobacteria from the oral cavity are able to colonize arterial plaque in individuals with heart disease?**
Application Questions
Enteric bacteria, lactic acid bacteria, and propionic acid bacteria have distinctive metabolic traits that can be used to characterize and identify these organisms. Describe the metabolic characteristics of these organisms, name a genus that belongs to each group, and indicate in what way these organisms can be differentiated.
Microorganisms have a variety of relationships with oxygen. Describe the terms used to characterize a cell’s response to oxygen and give an example from this chapter of an organism that can be described by each of these terms.
Chapter Glossary
a property of Mycobacterium species in which cells stained with the dye basic fuchsin resist decolorization with acidic alcohol Actinomycetes
a term used to refer to aerobic filamentous bacteria in the phylum Actinobacteria Coryneform bacteria
gram-positive, aerobic, nonmotile, rod-shaped organisms with the characteristic of forming irregular-shaped, club-shaped, or V-shaped cell arrangements, typical of several genera of unicellular Actinobacteria Enteric bacteria
a large group of gram-negative, rod-shaped Bacteria characterized by a facultatively aerobic metabolism and commonly found in the intestines of animals Heterofermentative
in reference to lactic acid bacteria, capable of making a mixture of fermentation products typically including lactate, ethanol and CO2 High G + C gram-positive bacteria
a term that refers to bacteria in the Actinobacteria Homofermentative
in reference to lactic acid bacteria, producing only lactic acid as a fermentation product Lactic acid bacteria
fermentative bacteria that produce lactic acid, are found in the Firmicutes, and are important in the production and preservation of many foods Low G + C gram-positive bacteria
a term that refers to bacteria in the Firmicutes Oligotrophic
a term that refers to organisms that grow best under low-nutrient conditions Propionic acid bacteria
gram-positive fermentative bacteria that generate propionate as a fermentation end product and are important in the production of cheese Proteobacteria
the largest and most metabolically diverse phylum of bacteria Pseudomonad
a term used to refer to any gram-negative, polarly flagellated, aerobic rod able to use a diverse suite of carbon sources