26 Innate Immunity: Broadly Specific Host Defenses
26 Innate Immunity: Broadly Specific Host Defenses
## Chapter 26 Innate Immunity: Broadly Specific Host Defenses
Periodontal Disease and Alzheimer’s: Evidence for Causation?
Alzheimer’s disease (AD) is characterized by the presence of insoluble aggregates of amyloid-β protein (Aβ) in the brain, producing plaques that interfere with cognitive brain function. Although Aβ has traditionally been viewed as an injurious substance, studies have now shown that it may function as an innate antimicrobial defense to protect the brain from bacteria that cross the blood–brain barrier. In such cases, the invading cells become entombed in a meshwork of Aβ, minimizing their growth and dissemination in the brain (see Figure 26.3). Chronic or heavy bacterial infections are believed to initiate the Aβ plaques that lead to Alzheimer’s symptoms, and a recent report has implicated a specific bacterium in the development of these plaques: Porphyromonas gingivalis, the causative agent of periodontal disease.
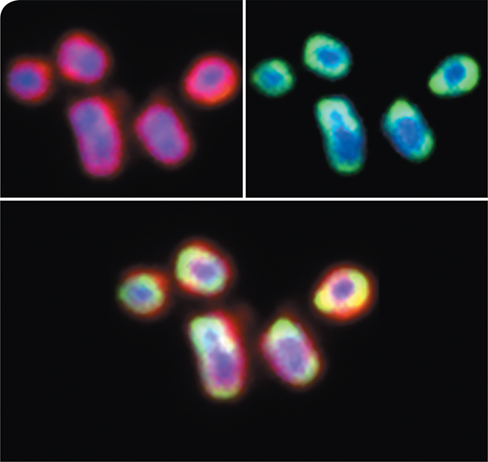
It has long been recognized that Alzheimer’s patients also often suffer from chronic gum disease, including gingivitis and periodontitis, and multiple reports are emerging that suggest the onset of gum disease is a risk factor that precipitates AD. In one study, researchers introduced oral P. gingivalis infection in mice and subsequently documented P. gingivalis colonization of the brain. There, the invading bacteria produced neurotoxic proteases called gingipains (red, on cells of P. gingivalis, top left photo). This triggered inflammation and accumulation of Aβ (green, on cells, top right photo), which colocalized with the gingipains on cells of P. gingivalis (bottom photo), ultimately promoting Aβ plaque formation. However, when gingipain inhibitors were administered, bacterial numbers diminished, inflammation and production of Aβ subsided, and neurons in the hippocampus were restored, suggesting that such inhibitors may reduce Aβ plaque formation and serve as a new weapon against AD.
Although the study is not conclusive, the results indicate that P. gingivalis infection may be a contributing factor in the development of AD. Further study along these lines may eventually yield a more complete understanding of AD pathology, as well as more effective treatment options.
Source: Dominy, S.S., et al. 2019. Porphyromonas gingivalis in Alzheimer’s disease brains: Evidence for disease causation and treatment with small-molecule inhibitors. Sci. Adv. 5. doi: 10.1126/sciadv.aau3333.
We began this unit by considering the associations of microbes with humans, and in the last chapter we discussed pathogenicity, virulence, and risk factors for infection. In this and the next chapter, we focus on mechanisms used by vertebrates to resist pathogens and the diseases they cause, the science of immunology. In this chapter, we present concepts of innate immunity, inborn host defenses against a broad range of pathogens. In Chapter 27 we discuss adaptive immunity, the essential second tier of the immune system that targets specific pathogens to minimize their harmful effects.
I Fundamentals of Host Defense
The human body has both innate (broadly targeted) and acquired (highly specific) immune mechanisms to either prevent or respond to invasion by pathogens. This highly developed “search and destroy” system maintains human health from potentially harmful microbes in the environment.
We begin with an overview of innate and adaptive immunity and follow this with a consideration of the human body’s significant natural barriers to invasion by pathogens.
26.1 Basic Properties of the Immune System
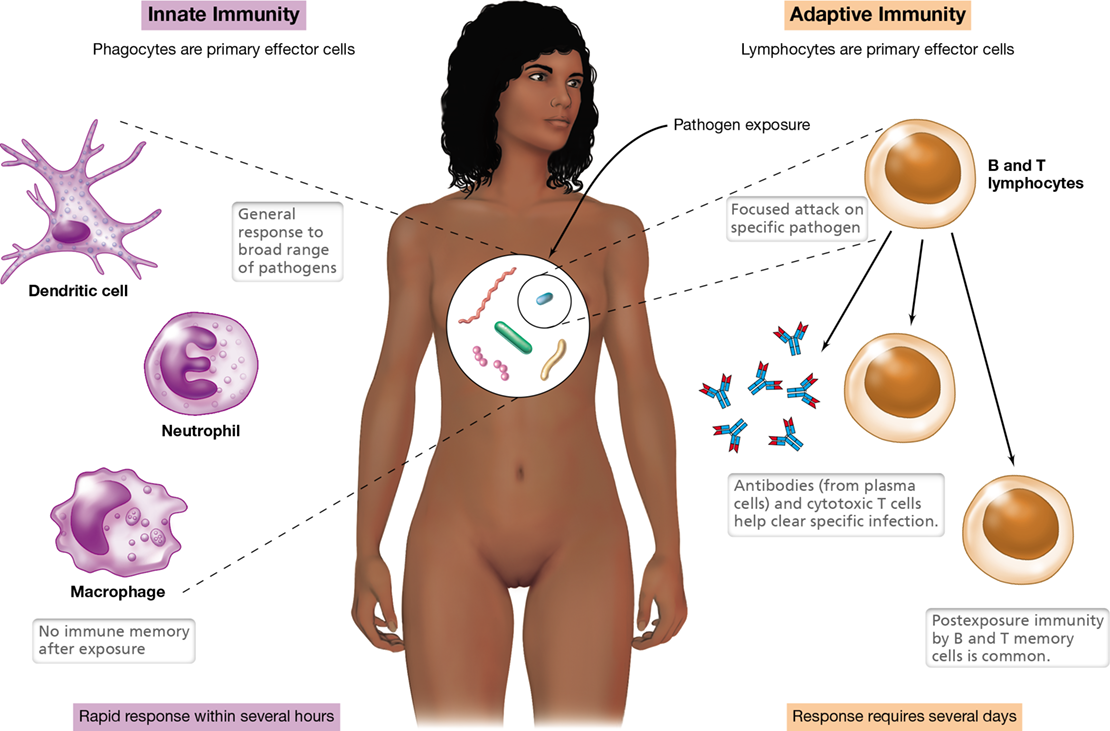
Immunity is the ability of an organism to resist infection. The human immune system employs a two-pronged defense against invading pathogens. Innate immunity, the first of these interconnected defensive mechanisms, is the built-in capacity of the immune system of multicellular organisms to target pathogens that are seeking to colonize the host. Depending on the virulence of the pathogen, the infectious dose, and the immune competence of the individual, innate mechanisms alone may be insufficient to eliminate the pathogen. In such cases, and assuming that the pathogen has not quickly killed the host, adaptive immunity is triggered within the individual against those specific pathogens that have overcome the innate defenses. Each adaptive immune response is specifically targeted to a particular invading pathogen, such as a single strain of virus. Figure 26.1 compares and contrasts the fundamental elements of each of these indispensable branches of the immune system.
Figure 26.1 Overview of the two-pronged immune response.

Principal characteristics of innate (shown in purple) and adaptive (shown in orange) immunity. Phagocytes and lymphocytes are the primary effector cells of innate and adaptive immunity, respectively, but note that not all immune cells are shown (compare to Figure 26.5).
Mastering Microbiology
Art Activity: Figure 26.1 Overview of the two-pronged immune response
Principles of Innate Immunity
Innate immunity is a noninducible, preexisting ability of the body to recognize and destroy a broad range of pathogens or their products. Innate immune responses develop quickly, typically within several hours of exposure to a pathogen or its product, and they do not require previous exposure to a pathogen or its products for activation. Eukaryotes have functionally similar pathogen recognition mechanisms that lead to rapid and effective host defense. For example, pathogen recognition in the insect Drosophila (fruit fly) is carried out in much the same way as it is in humans, using immune cells that show structural and evolutionary homology.
In addition to a variety of physical and chemical barriers to infection that we will describe in Section 26.2, innate immunity is largely dependent on the activity of phagocytes (Figure 26.1), cells that ingest, kill, and digest microbial pathogens. Phagocytes include neutrophils, macrophages, dendritic cells, and eosinophils, and we define their distinctive features in Section 26.4. In addition to phagocytes, cells of the innate immune system consist of mast cells and basophils, both of which trigger inflammation when activated (Section 26.8), and natural killer (NK) cells, which identify and destroy host cells that have become compromised and are therefore dangerous to the host, such as cells that are infected by a pathogen or are cancerous (Section 26.10).
Mastering Microbiology
Art Activity: Figure 26.2 Barriers to infection in the human body
Principles of Adaptive Immunity
When innate immune responses fail to eliminate an invading pathogen, adaptive immunity is activated. Adaptive immunity is the acquired ability to recognize and destroy a specific pathogen or its products. Adaptive responses show specificity because they are directed at unique pathogen components or products called antigens, which often define a particular strain of pathogen or type of foreign material. Macrophages and dendritic cells ingest and process pathogens to present peptide antigens to T lymphocytes (T cells), which play a key role in initiating the adaptive response. Another adaptive immune cell, the B lymphocyte (B cell), also presents antigen to T cells. The presented antigen peptides, called epitopes, bind specific receptors on the lymphocyte surface, triggering the expression of genes that cause the lymphocytes to proliferate and differentiate. Differentiated B cells called plasma cells specialize in the production of antigen-specific proteins called antibodies (immunoglobulins), which bind the pathogen and mark it for destruction.
Unlike the comparatively rapid innate response, a protective adaptive response usually takes several days to develop, and the strength of the adaptive response increases as the numbers of antigen-reactive lymphocytes increase. Although adaptive immune responses are typically slower than innate response mechanisms, they are highly specific and result in immune memory, the ability of lymphocytes to quickly respond after subsequent exposure to a previously encountered antigen.
With this broad overview of inborn and acquired immune responses in place, we now consider the many natural barriers to infection that exist in the healthy human body. It is only after one or more of these barriers has been breached that pathogen invasion can occur and the immune system is brought into play.
Check Your Understanding
What major class of immune cells mediates an innate immune response? What additional type of immune cells is required for an adaptive immune response?
What term is used to describe the unique molecules found on the surface of different pathogens?
26.2 Barriers to Pathogen Invasion
Although the vast majority of microorganisms that associate with the human body are not pathogenic, certainly some do cause disease. The best defense against infectious diseases for the human body is to prevent pathogens from colonizing in the first place. Humans have a host of resistance factors that inhibit pathogen colonization and thereby prevent the onset of infectious diseases. These include a variety of physical and chemical barriers to microbial infection. In addition, the condition of the host and the composition of microorganisms that normally inhabit the host are factors that are often the tipping point between health and disease.
Natural Host Resistance
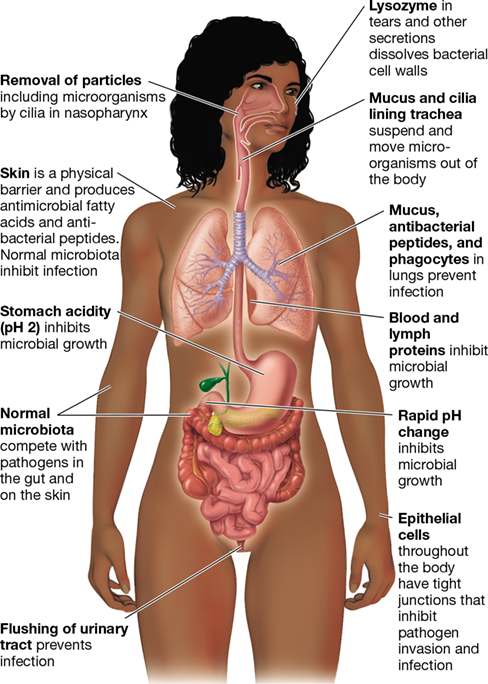
Several resistance factors common to vertebrate hosts inhibit infection by most pathogens in a nonspecific way (Figure 26.2). For example, the presence of normal microbiota in the human body is critically important for resisting pathogen infection, especially on the skin and in the mucosal tissues of the gastrointestinal, respiratory, and reproductive tracts (Chapter 24). Pathogens do not easily infect tissues on which normal microbiota are well established because the harmless microbes limit available nutrients and sites for infection by the pathogens. Although the competitive exclusion of invading pathogens by resident microbes is technically not a component of the host’s immune system, the nonpathogenic normal microbiota found in or on the body play a major role in preventing disease through this principle. It is not uncommon for disruptions to the composition of the normal microbiota, such as that which may occur when antibiotic drug treatments nonspecifically kill harmless and even beneficial microbes in the body, to trigger the onset of disease by opportunistic pathogens.
Figure 26.2 Barriers to infection in the human body.

These barriers provide natural resistance to colonization and infection by pathogens.
The ability of a particular pathogen to cause disease in a host is highly variable. For instance, certain animal species, including raccoons and skunks, are much more susceptible to the rabies virus than are others, such as the opossum, which only rarely develops the disease. Anthrax infects many species of animals, causing disease symptoms varying from fatal blood poisoning in cattle to the mild pustules of human cutaneous anthrax. Introduction of the same pathogen by other routes, however, may challenge the resistance of the host. For example, anthrax causes a localized infection when acquired through the skin but a lethal, systemic infection when acquired through the mucous membranes of the lungs (Sections 30.9 and 32.8). Another barrier to infection is the fact that diseases of endothermic (“warm-blooded”) animals, such as birds and mammals, are rarely transmitted to ectothermic (“cold-blooded”) animals, such as amphibians and reptiles, and vice versa. Presumably, the anatomical and physiological features of one group are not compatible with pathogens that infect the other group.
In addition to these factors, the general condition of the host plays a role in the balance between health and disease. Susceptibility to infections is higher in the very young and the very old, as well as those suffering from nutritional or stress-related problems. For example, the intestinal microbiota matures over time and thus infants under one year of age often contract diarrhea caused by bacterial or viral pathogens more readily than do adults. Conversely, the immune system of older adults may be weakened or compromised from medical procedures or drug therapy, leading to a higher susceptibility to infectious disease. Other problems that affect both the young and the old—smoking, poor diet, intravenous drug use, excessive alcohol consumption, chronic lack of sleep—can all play a role in whether a pathogen can infect a host and cause disease.
Infection Site and Tissue Specificity
Most pathogens must adhere to and colonize the site of exposure to initiate infection. Even if pathogens adhere to an exposure site, the organisms cannot colonize the host if the site is not compatible with the pathogen’s nutritional and metabolic needs. For example, if cells of Clostridium tetani (cause of the disease tetanus) were ingested, tetanus would not normally result because the pathogen would be either killed by the acidity of the stomach or unable to compete with the well-developed normal intestinal microbiota. If, on the other hand, C. tetani cells or endospores were introduced into a puncture wound, the organism would grow and produce tetanus toxin in the anoxic zones created by local tissue death. Conversely, enteric bacteria, such as Salmonella and Shigella, do not normally cause wound infections but can infect the intestinal tract and cause gastrointestinal illness.
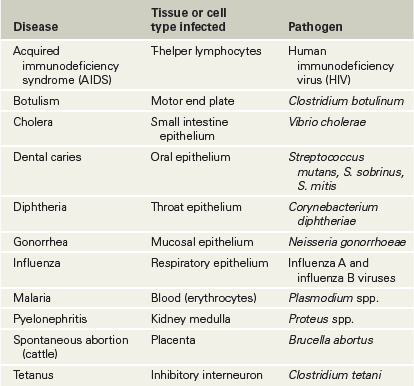
In some cases, pathogens interact exclusively with members of a few closely related host species because the hosts share tissue-specific receptors, important in establishing infection (Section 25.1). For example, human immunodeficiency virus (HIV, the causative agent of AIDS) infects only humans and their closest primate relatives, including the great apes. This is because the HIV-binding cell surface proteins CXCR4, present on human T lymphocytes, and CCR5, present on human macrophages (Section 26.1), are also expressed in great apes. Other animals, including most other primates, lack CXCR4 and CCR5 and are therefore not susceptible to HIV infection. Many other pathogens exhibit tissue specificity to such a degree that they infect only a single species. For example, a rhinovirus that infects nasal epithelial cells in humans, causing symptoms of the common cold, would generally not infect and cause similar symptoms in a nonhuman host. Table 26.1 presents some other examples of tissue specificity in pathogens.
Table 26.1 Tissue specificity in infectious disease

Physical and Chemical Barriers to Infection
The structural integrity of tissue surfaces poses a barrier to penetration by microorganisms (Section 25.2). The tight junctions between epithelial cells that line body tissues inhibit invasion and infection (Figure 26.2). In the skin and mucosal tissues, potential pathogens must first adhere to tissue surfaces and then grow at these sites before traveling elsewhere in the body. Mucosal surfaces are coated with a layer of protective mucus that traps microorganisms, pollen, and other foreign agents. Epithelial cells underlying the mucus layer have cilia on their surfaces that carry out coordinated movements to expel suspended pathogens and keep them from adhering to tissues. This combination of mucus and ciliary motion plays a key role in removing inhaled microorganisms from the mucosal surfaces of the trachea and bronchial tubes, thus preventing microbial colonization of the lungs.
In the central nervous system (CNS), pathogens that are able to penetrate the blood–brain barrier present a special challenge because adaptive immune responses mediated by lymphocytes are less effective at these sites. However, a long-misunderstood protein—amyloid-β (Aβ)—may play a role in innate defense of the brain by containing invading pathogens. As discussed in MicrobiologyNow at the beginning of the chapter and shown in Figure 26.3, Aβ proteins form a mesh of fibrils to trap bacteria that are able to invade the CNS. However, in cases of heavy or chronic infection, excessive deposition of Aβ may cause the fibrils to aggregate into dense, insoluble complexes that form amyloid plaques in the brain tissue, one of the hallmarks of Alzheimer’s disease.
Figure 26.3 Amyloid-β (Aβ) protein as an innate defense in the central nervous system.

(a) Bacteria that breach the blood–brain barrier stimulate production of Aβ protein, which forms a dense network of fibrils to trap the pathogens and minimize their invasiveness. (b) Fluorescence micrograph of cells of Salmonella bacteria (green) immobilized by a mesh of Aβ fibrils (red), enabling more efficient pathogen destruction by immune mechanisms.
Sebaceous glands in the skin secrete fatty acids and lactic acid, lowering the acidity of the skin to pH 5 and inhibiting colonization by many pathogenic bacteria (blood and most internal organs are about pH 7.4). Potential pathogens ingested in food or water must survive the strong acidity (pH 2) and digestive enzymes, such as pepsin, in the stomach (Figure 26.2). Although the pH returns to neutral in the lower intestinal tract, pathogens that survive passage through the stomach must then compete with the abundant resident microbiota present in the small and large intestines (Figure 24.3). In addition to the established normal microbiota that exist at many potential infection sites, resistance to infection and invasion is enhanced by antibacterial substances called defensins, produced in the skin, lungs, and gut. Finally, the lumen of the kidney, the eye, the respiratory system, and the cervical mucosa are constantly bathed with tears, mucus, or other secretions that contain lysozyme, an enzyme that can kill bacteria by digesting the cell wall.
Mastering Microbiology
Art Activity: Figure 26.3a The blood and lymphatic systems
The human body is thus naturally protected by a host of physical barriers, chemicals, and secretions, all of which combine to naturally suppress pathogen invasion and infection. However, breaching of these defenses by a pathogen activates cellular defenses of the immune system, beginning with mechanisms of microbe recognition and innate responses. We now consider the organs and cellular components of the immune system before exploring how this complex and interrelated system works.
Check Your Understanding
How do pathogens recognize the tissues they infect?
Identify physical and chemical barriers to pathogens. How might these barriers be compromised?
What other factors may control the outcome of an infectious disease?
II Cells and Organs of the Immune System
Cellular components of the immune system are derived from hematopoietic stem cells in the bone marrow. These cells circulate throughout the body in the blood and lymph systems in constant surveillance for foreign cells, viruses, or their antigenic products.
The immune response is a result of the activities of a wide variety of specialized cells that circulate throughout the body, primarily through the blood and lymph, a fluid similar to blood but which lacks red blood cells.
26.3 The Blood and Lymphatic Systems
Circulatory systems in the human body include separate vasculature for blood and lymph. It is through circulation of these fluids that cells and proteins of the immune system are transported to the various tissues and organs of the body. All cells found in the blood and lymph are derived from hematopoietic stem cells found primarily in the bone marrow but also in the gut. Hematopoietic stem cells continuously divide and differentiate to supply the body with both erythrocytes (red blood cells) and leukocytes (white blood cells).
Blood and Lymph Circulation
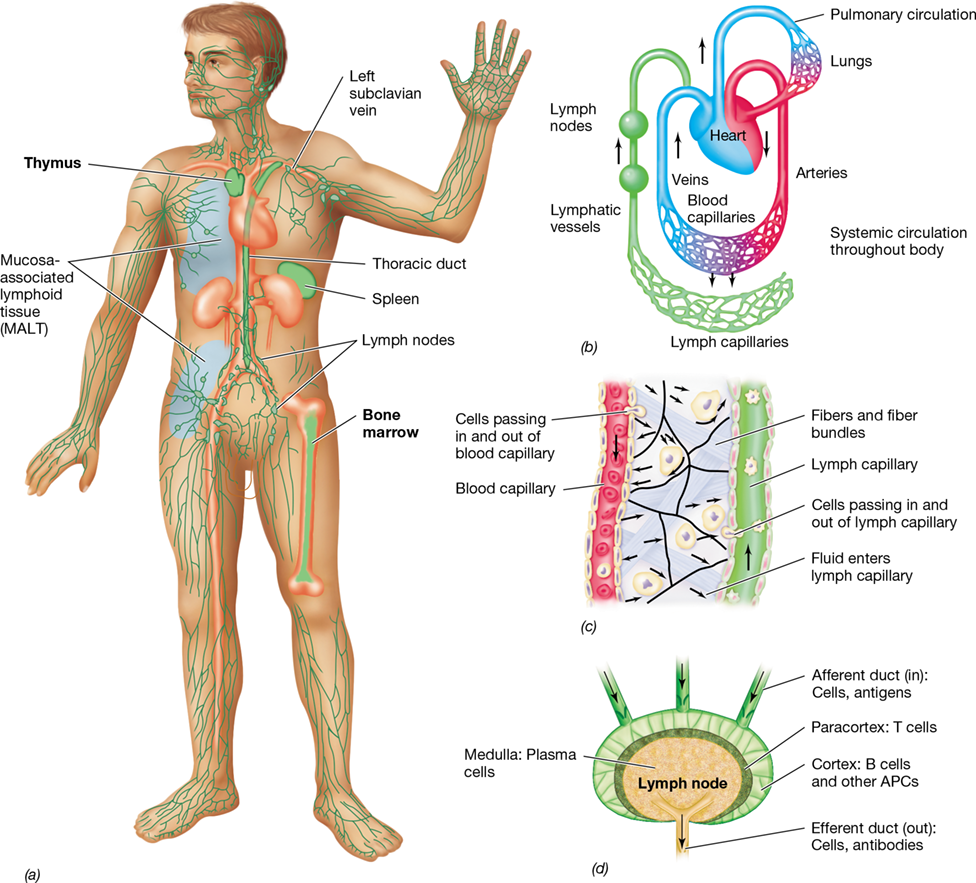
Blood is pumped by the heart through arteries and capillaries throughout the body and is returned to the heart through the veins (**Figure 26.4a,*b***). In the capillary beds, leukocytes and solutes pass to and from the blood into the lymphatic system. Lymph drains from extravascular tissues into lymphatic vessels (lymph capillaries and lymph ducts) and then into lymph nodes throughout the lymphatic system (Figure 26.4b–d). Lymph nodes contain leukocytes arranged to encounter antigens, such as those found on microorganisms, as they travel through the lymphatic circulation. Lymph containing antibodies and immune cells empties into the blood circulatory system via the thoracic lymph duct (Figure 26.4a). Mucosa-associated lymphoid tissue (MALT) is another organized lymphoid tissue containing leukocytes that interact with antigens that enter the body through mucous membranes, including those of the gut, the genitourinary tract, and the respiratory tract.
Figure 26.4 The blood and lymphatic systems.

(a) Blood and lymph circulation in the body. Major blood vessels and associated organs are shown in red. Major lymphatic organs and vessels are shown in green. The primary lymphoid organs are the bone marrow and thymus. The secondary lymphoid organs are the lymph nodes, spleen, and mucosa-associated lymphoid tissue (MALT). (b) Connections between the lymphatic and blood systems. Blood flows from the veins to the heart, to the lungs, and then through the arteries to capillaries in tissues. Exchange of solutes and cells occurs between blood and lymphatic capillaries. Lymph drains from the thoracic duct into the left subclavian vein of the blood circulatory system. (c) The exchange of cells between the blood and lymphatic systems is shown in detail. Both blood and lymphatic capillaries are closed vessels, but cells pass from blood capillaries to lymphatic capillaries and back by the process of diapedesis. (d) A secondary lymphoid organ, the lymph node, showing the major anatomical areas and the immune cells concentrated in each area. The anatomy of the MALT and the spleen is analogous to that of the lymph nodes. APCs, antigen-presenting cells.
The spleen is an important organ in immunity and consists of red pulp—rich in red blood cells and macrophages—and white pulp, which consists of organized lymphocytes and phagocytes arranged to filter blood in a manner similar to lymph nodes and MALT. Collectively, the lymph nodes, MALT, and spleen are called secondary lymphoid organs (or tissues, in the case of MALT, Figure 26.4). The secondary lymphoid organs are the sites where antigens interact with antigen-presenting cells (Section 26.4) and lymphocytes to trigger the adaptive immune response.
Hematopoiesis
Hematopoietic stem cells are precursor cells found in bone marrow and the gut that can differentiate into any type of blood cell (see Figure 26.5), a process called hematopoiesis. Stem cell differentiation in these tissues is influenced by soluble cytokines and chemokines, proteins that direct immune cell production, function, and movement, thereby modulating the immune response (Section 26.5). Following differentiation, newly formed blood cells enter the circulation and reach other parts of the body, where the leukocytes further mature and become activated upon antigen encounter.
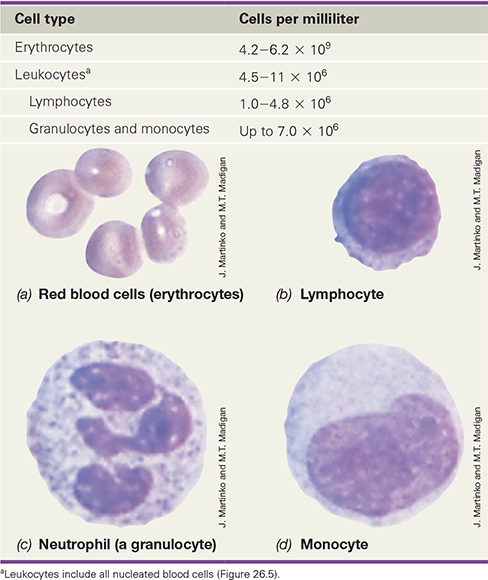
Blood consists of cellular and noncellular components, including many cells and molecules active in the immune response (Table 26.2). The most numerous cells in human blood are erythrocytes, small, nonnucleated cells that carry oxygen from the lungs to the tissues. About 0.1% of the cells in blood, however, are white blood cells—leukocytes—of which there are many different types (Table 26.2). Blood cells are suspended in plasma, a liquid that contains proteins and other solutes. Outside the body, blood or plasma quickly forms an insoluble fibrin clot, remaining liquid only when an anticoagulant such as potassium citrate or heparin is added. When blood clots, the insoluble proteins trap the cells in a large, insoluble mass. The remaining fluid, called serum, lacks both cells and clotting proteins. Serum does, however, contain a high concentration of other proteins, in particular antibodies, key players in the adaptive immune response, as we will learn in Chapter 27.
Table 26.2 Major cell types found in normal human blood

Check Your Understanding
Describe the circulation of a leukocyte from the blood to the lymph and back to the blood.
What soluble molecules determine whether a particular hematopoietic stem cell will become a phagocyte, lymphocyte, or erythrocyte?
26.4 Leukocyte Production and Diversity
The innate and adaptive immune responses are mediated by several types of leukocytes (Table 26.2). From its hematopoietic stem cell precursor, a leukocyte will differentiate and mature through either the myeloid or the lymphoid lineage (Figure 26.5). Leukocytes move throughout the body and, through a process called diapedesis (also called extravasation), pass from blood to interstitial spaces. Leukocytes may then be collected with lymph into lymphatic vessels, where they are eventually returned to the blood circulatory system (Figure 26.4c).
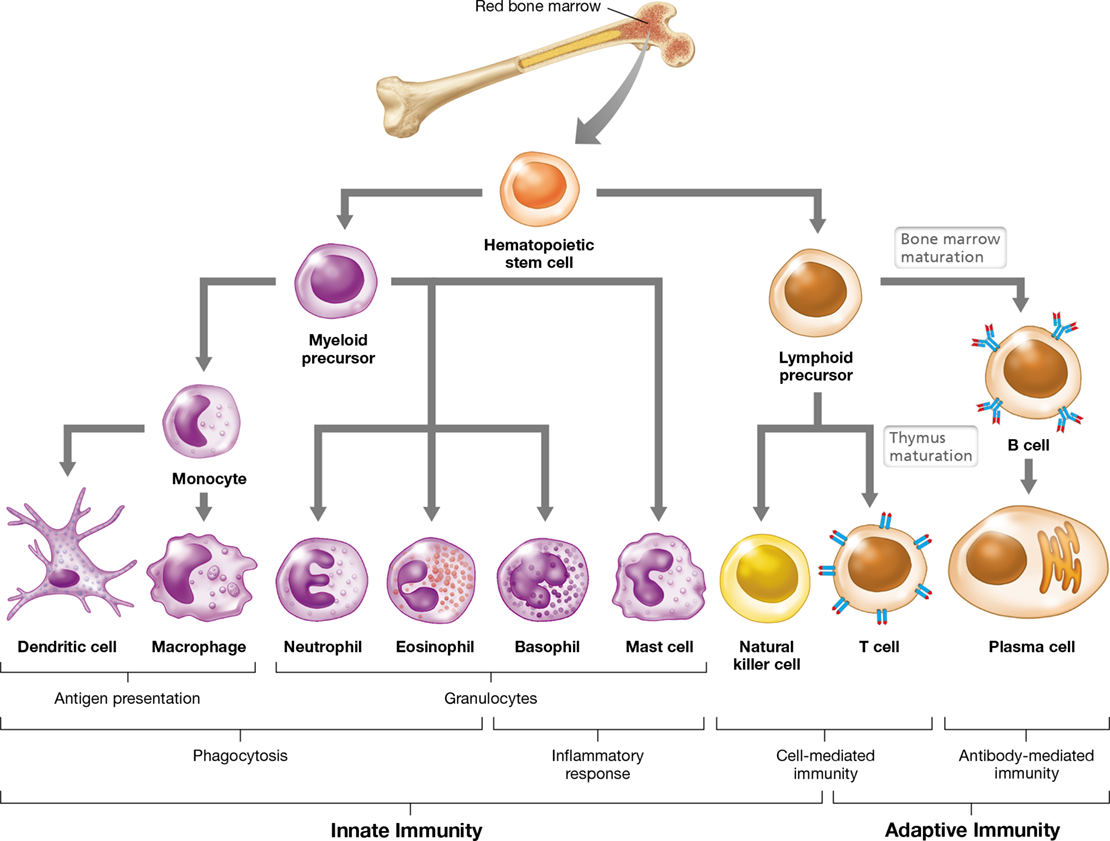
Figure 26.5 Lineage and diversity of immune response cells.

Immune cells develop from hematopoietic stem cells in the bone marrow into either myeloid precursors or lymphoid precursors. These precursors, in turn, differentiate into mature cells that have various immune functions. (Erythrocytes also develop from myeloid precursors.)
Myeloid Cells—Monocytes and Granulocytes
Myeloid cells, the principal effector cells of innate immunity, are derived from myeloid precursor cells. Mature myeloid cells develop from one of two lineages: monocytes or granulocytes (Figure 26.5 and Table 26.2). Immature monocyte precursor cells circulate in the blood for several days before moving into other tissues and differentiating into macrophages or dendritic cells. These phagocytic cells function as antigen-presenting cells (APCs) because they use a special type of cell surface protein called a class II major histocompatibility complex (MHC) to present engulfed and processed antigens to T lymphocytes, which then initiate an adaptive immune response (Figure 26.5). We will revisit MHC proteins later in this chapter and discuss them in detail in Chapter 27.
Macrophages are often the first defensive cells that interact with a pathogen and are abundant in many tissues and organs, especially the spleen, lymph nodes, and MALT, where they constitute up to 15% of total cells. Because they ingest and destroy most pathogens and foreign molecules that invade the body, macrophages are essential to the innate response. Dendritic cells are found throughout the body tissues and are especially abundant along epithelial linings, including the skin and mucous membranes. When dendritic cells ingest antigen, they migrate to nearby lymph nodes (Figure 26.4), where they are extremely efficient at presenting antigen to T lymphocytes. Thus, dendritic cells are an important link between innate and adaptive immunity. The specialized antigen-presenting properties of macrophages and dendritic cells will be examined in Section 27.5.
Granulocytes are the second lineage of cells derived from myeloid precursors. Granulocytes contain cytoplasmic inclusions, or granules, that are visible in stained microscopic preparations. These granules contain toxins or enzymes that are released to destroy target cells. One granulocyte, the neutrophil, also called a polymorphonuclear leukocyte (PMN), is an abundant, highly motile phagocyte that responds rapidly to pathogen challenge, an activity that is central to innate immunity. Neutrophils are found predominantly in the bloodstream and bone marrow, from where they migrate to sites of active infection. Eosinophils are a second, much less numerous, type of phagocytic granulocyte. A major function of eosinophils is to destroy larger, extracellular parasites, such as infectious helminths (worms, Section 34.7), by secreting cytotoxic enzymes and then clearing the debris from the body tissues.
Other granulocytes include basophils, which circulate in the blood, and mast cells, which reside in host tissues (Figure 26.5). The intracytoplasmic granules of these cells contain histamine and other mediators that function to initiate an inflammatory response when secreted, a process called degranulation. Mast cell and basophil degranulation is responsible for certain types of allergic reactions (Section 28.1).
Lymphocytes
Lymphocytes are specialized leukocytes derived from lymphoid precursor cells (Figure 26.5 and Table 26.2). As we have discussed (Section 26.1), there are two major types of lymphocytes: B cells (B lymphocytes) and T cells (T lymphocytes) (Figure 26.5). Whereas B cells originate and mature in the bone marrow, T cells begin their development in the bone marrow but migrate to the thymus to mature (Figure 26.4a and see Figure 27.2). In mammals, the bone marrow and thymus are called primary lymphoid organs because they are the sites where lymphoid stem cells differentiate into functional, antigen-specific lymphocytes (Figure 26.4a). Naive B and T cells circulate through the blood and lymph system and into the secondary lymphoid organs (Section 26.3) where, upon antigen encounter, they become activated and further differentiate into effector and memory cells.
B cell surfaces are covered with B cell receptors (BCRs), membrane-bound immunoglobulins that function to bind free antigen. Following internalization and processing of bound antigen, B cells—like dendritic cells and macrophages—function as specialized APCs, employing class II MHCs to present antigen to T lymphocytes (see Figure 27.6). The T cells, in turn, interact with presented antigens through membrane-bound T cell receptors (TCRs), protein complexes that specifically bind antigens presented on MHCs. We explore the roles of B and T lymphocytes in the adaptive immune response in Chapter 27.
Like B and T cells, natural killer (NK) cells (Figure 26.5) are derived from lymphoid precursors. However, unlike B and T lymphocytes, NK cells function primarily in innate immunity. NK cells rid the body of virus-infected and tumor cells using a mechanism that distinguishes healthy versus compromised cells based on the presence (or absence) of specific surface molecules (Section 26.10).
Check Your Understanding
How does the development of B, T, and NK cells differ from that of dendritic cells and macrophages?
Distinguish between the primary lymphoid organs and the secondary lymphoid organs. What is the relationship between the components of these two groups?
III Phagocyte Response Mechanisms
Phagocytes respond quickly to tissue damage and microbial invasion, whereupon they recognize, engulf, and digest pathogens through various molecular mechanisms. Phagocytic activities are the hallmark of the innate immune response.
Innate immunity is primarily driven by the activities of phagocytes (Section 26.1). These cells recognize common structural features found on and in pathogens. Interactions with pathogens activate genes in the phagocytes that control the transcription, translation, and expression of proteins that eventually lead to the destruction of the pathogens.
Innate immunity responds within minutes of a phagocyte engaging a pathogen, but it is not always effective in controlling infections. To counter these infections, certain phagocytes also activate adaptive immunity by processing and presenting antigens to receptors on T lymphocytes (Chapter 27). However, because an adaptive response to a specific pathogen takes several days to develop, the ability of phagocytes to quickly respond to microbial invasion is an indispensable mechanism for infection containment.
26.5 Pathogen Challenge and Phagocyte Recruitment
When a pathogen breaches the physical and chemical barriers of a host (Figure 26.2) and causes an infection, the immune system is mobilized to protect the host from further damage. Innate immunity is the first line of defense and is critical for host protection for about four days after an infection begins. Phagocytes engulf and destroy pathogens, often initiating complex host-mediated inflammatory reactions when they do (Section 26.8).
Microbial Invasion
The initial inoculum of a pathogen is usually insufficient to cause host damage, even if a pathogen gains access to tissues. For the pathogen to be successful, it must attach, multiply, and colonize the tissue (Figure 25.1), and these events require that the pathogen find appropriate nutrients and environmental conditions for growth.
Following colonization, a pathogen must usually invade tissues to initiate disease. Invasion is the ability of a pathogen to enter into host cells or tissues, multiply, spread, and cause disease. Microbial infections often begin at breaks or wounds in the skin (**Figure 26.6*a***) or on the mucous membranes of the respiratory, digestive, or genitourinary tract, surfaces that are normally microbial barriers (Figure 26.2). In some cases, growth may also begin on intact mucosal surfaces, especially if the composition of the normal microbiota has been altered or eliminated, for example by antibiotic therapy. Tissue damage caused by invasive pathogens or injury triggers the recruitment of large numbers of phagocytes and other immune cells to the site of infection.
Figure 26.6 Microbial invasion and the innate immune response.

(a) Tissue damage, such as that caused by a tack puncturing the skin, can lead to invasion by microorganisms and the release of cytokines and chemokines from damaged host cells. (b) Phagocytes, especially neutrophils, are recruited to the site of infection by the cytokine–chemokine gradient, squeezing out of dilated capillaries via diapedesis. (c) Invading microorganisms are cleared by phagocytosis, and the tissue is restored to health. See Figures 26.1 and 26.5 for functional descriptions of some of the cells and components shown here, and see Table 26.1 for photomicrographs and quantitative data on some of the cells shown here.
Tissue Damage and Chemokine Release
When microorganisms invade and damage host tissues, resident phagocytes (usually tissue macrophages) engage the microbes and become activated. The activated macrophages and damaged host cells release cytokines and chemokines into the surrounding tissues (Figure 26.6a). Cytokines bind specific receptors on cells and induce a particular response from them, usually by activating a signaling pathway that controls transcription and protein synthesis. Chemokines are an important subclass of cytokines that have the specific role of recruiting circulating immune cells, especially neutrophils (Figure 26.5), to the site of injury.
Resident macrophages, which are found in all of the body’s organ systems, are stimulated by the presence of invading pathogens to secrete chemokines that establish a gradient of chemoattractants. These molecules bind receptors on other immune cells, especially neutrophils and T cells, and recruit them to the site of infection (Figure 26.6b). This triggers a neutrophil-mediated inflammatory response and, later, an adaptive immune response facilitated by lymphocytes against the specific invading agent or its toxic products.
Localized inflammation allows neutrophils, the most numerous circulating phagocytes, to migrate quickly along the chemotactic gradient to the site of infection (Figure 26.6b). Once there, chemokine-mediated binding reactions promote neutrophil adhesion to the inner wall of the blood capillary, a process called margination. The halted neutrophils then undergo diapedesis, the movement of leukocytes from the bloodstream to surrounding infected tissues by squeezing between endothelial cells lining the capillaries (Figure 26.6b). Higher than normal numbers of neutrophils in the blood or at a site of inflammation indicate an active response to a current infection.
Neutrophils and other phagocytes that encounter pathogens in damaged areas must be able to recognize, capture, and destroy pathogens to clear infections and restore body tissues to a healthy state (Figure 26.6c). We explore the molecular mechanisms that facilitate these cellular interactions in the next section.
Check Your Understanding
Describe several mechanisms by which opportunistic microorganisms can invade body tissues. What body sites are often associated with microbial invasion?
Describe the mechanisms by which circulating phagocytic cells are recruited to a site of infection.
26.6 Pathogen Recognition and Phagocyte Signal Transduction
26.6 Pathogen Recognition and Phagocyte Signal Transduction
26.6 Pathogen Recognition and Phagocyte Signal Transduction
The first type of immune cell to be activated in the innate response is typically a phagocyte, whose primary function is to engulf and destroy pathogens. Phagocytes include neutrophils, eosinophils, and monocytes, which are found primarily in the blood, and macrophages and dendritic cells, which occur primarily in the body tissues (Figure 26.5). But how do these cells distinguish pathogens and other foreign agents from the myriad other cells in the body? Moreover, once invaders are recognized, how do phagocytes capture and destroy them? We address these topics now.
Pathogen-Associated Molecular Patterns
The macromolecules inside and on the surface of pathogens contain pathogen-associated molecular patterns (PAMPs), repeating structural subunits common to broadly related groups of infectious agents. PAMPs may include polysaccharides, proteins, nucleic acids, or even lipids. For example, a common PAMP is the lipopolysaccharide (LPS) common to all gram-negative bacterial outer membranes (Section 2.4). Other PAMPs include bacterial flagellin, the double-stranded RNA (dsRNA) of certain viruses, and the lipoteichoic acids of gram-positive bacteria (Section 2.3). Like all PAMPs, these molecules are found on various pathogens but are absent from host cells. Therefore, PAMPs serve as markers by which phagocytes can identify microbes, even if the pathogens have not been encountered previously.
Pattern Recognition Receptors
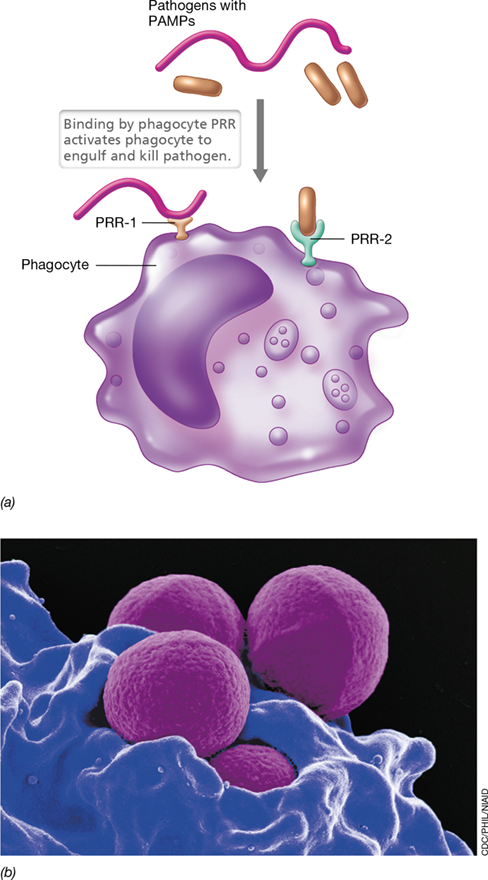
Phagocytes employ a pathogen-recognition system that triggers a timely and appropriate response, leading to recognition, containment, and destruction of pathogens. Phagocytes produce soluble or membrane-bound proteins called pattern recognition receptors (PRRs) that recognize and bind PAMPs, allowing phagocytes to interact quickly and effectively with pathogens (**Figure 26.7*a***). The interaction of a PAMP with a PRR activates the phagocyte to ingest and destroy the targeted pathogen by phagocytosis (Figure 26.7b and Section 26.7).
Figure 26.7 Pathogen recognition by phagocytes.

(a) Phagocytes interact with pathogens by way of preformed pattern recognition receptors (PRRs) that bind to pathogen-associated molecular patterns (PAMPs). Binding of a PAMP by a phagocyte PRR stimulates the phagocyte to destroy the pathogen and activate other phagocytes. (b) Colorized scanning electron micrograph of a neutrophil (blue) phagocytosing several cells of methicillin-resistant Staphylococcus aureus (MRSA; magenta). For more on MRSA, see Explore the Microbial World in Chapter 29 and Section 31.9.
PRRs were first observed in phagocytes of Drosophila, the fruit fly, where they are called Toll receptors, and they have since been identified as important components of microbial recognition throughout the animal kingdom (see Explore the Microbial World, “Pattern Recognition Receptors of Hydrothermal Vent Tube Worms Facilitate Endosymbiosis”). Structural, functional, and evolutionary homologs of the Toll receptors, called Toll-like receptors (TLRs), are widely expressed on mammalian innate immune cells. TLRs are found associated with membranes on the surface of or in intracellular vesicles of all types of phagocytes. At least nine TLRs in humans interact with a variety of cell surface and soluble PAMPs from viruses, bacteria, and fungi (Table 26.3).
Table 26.3 Receptors and targets in the innate immune response

aThe extracellular soluble PRRs are produced by liver cells in response to inflammatory cytokines.
bToll-like receptors initiate phagocyte activation via signal transduction.
cTLR-3, -7, -8, and -9 are found in membranes of intracellular organelles (endosomes) such as lysosomes. A tenth Toll-like receptor, TLR-10, has unknown ligand specificity and function.
dPresent in Baltimore class III viruses (Figure 11.2b and Section 11.10)
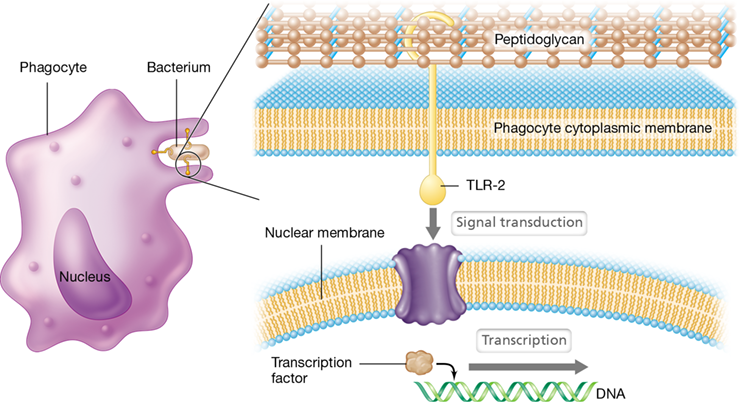
Each TLR on a human phagocyte recognizes and interacts with a specific PAMP. For example, TLR-2, a PRR on human phagocytes, interacts with peptidoglycan, a PAMP present in the cell wall of nearly all bacteria (Figure 26.8). This PAMP–PRR interaction activates the phagocytes, which then target gram-positive pathogens with exposed peptidoglycan. Access to the peptidoglycan of gram-negative bacterial cell walls is blocked by the surface lipopolysaccharides. However, another PRR found on phagocytes, TLR-4, interacts with the LPS of gram-negative bacteria, such as Salmonella spp., Escherichia coli, and Shigella spp. (Table 26.3 and see Figure 26.9).
Figure 26.8 A Toll-like receptor.
Membrane-spanning TLR-2 interacts with peptidoglycan from gram-positive pathogens. This interaction stimulates signal transduction (see Figure 26.9), activating transcription factors in the nucleus. The result is transcription of genes encoding proteins that induce inflammation and other phagocyte activities. All Toll-like receptors have analogous mechanisms for activating innate immunity.
Several soluble host molecules function similarly to these phagocyte-associated PRRs. The NOD-like receptors (NLRs) are a family of soluble PRRs found in the cytoplasm that have a nucleotide-binding domain (NOD). NOD1 and NOD2 interact with peptidoglycan components of gram-negative and gram-positive bacterial cell walls, respectively, stimulating production of antimicrobial peptides and inflammatory cytokines (Table 26.3). NOD-like receptor pyrin 3 (NLRP3) interacts with other proteins to form a structure called an inflammasome. Activation of the cytoplasmic inflammasome is mediated by cellular stress indicators, such as the loss of potassium ions (K+) from damaged cells and, especially, the binding of oxidized mitochondrial DNA produced in response to LPS and released into the cytosol; the latter triggers the production of proinflammatory cytokines, initiating inflammation. Later in this chapter we discuss the soluble PRRs in the context of their ability to activate proteins that enhance phagocytosis and destruction of pathogens (Section 26.9).
Signal Transduction in Phagocytes
Interaction of a PAMP with a PRR triggers transmembrane signal transduction, initiating gene transcription and translation of host-response proteins in a fashion similar to the two-component regulatory systems previously discussed in Bacteria and Archaea (Section 7.5). Signal transduction initiated by PAMP–PRR interaction results in enhanced phagocytosis, killing of pathogens, inflammation, and tissue healing.
For example, the binding of LPS (a PAMP derived from the cell wall of degraded gram-negative bacteria) to TLR-4 (a PRR on the surface of phagocytes) typically activates a signal transduction pathway (Figure 26.9). TLR-4 then binds proteins in the cytosol of the phagocyte, starting a cascade of reactions that activates transcription factors such as NFκB (nuclear factor kappa B), a protein that binds to specific regulatory sites on DNA, initiating transcription of downstream genes. Many of the NFκB-regulated genes encode host-response proteins, such as the cytokines that activate cells and initiate inflammation (Section 26.5).
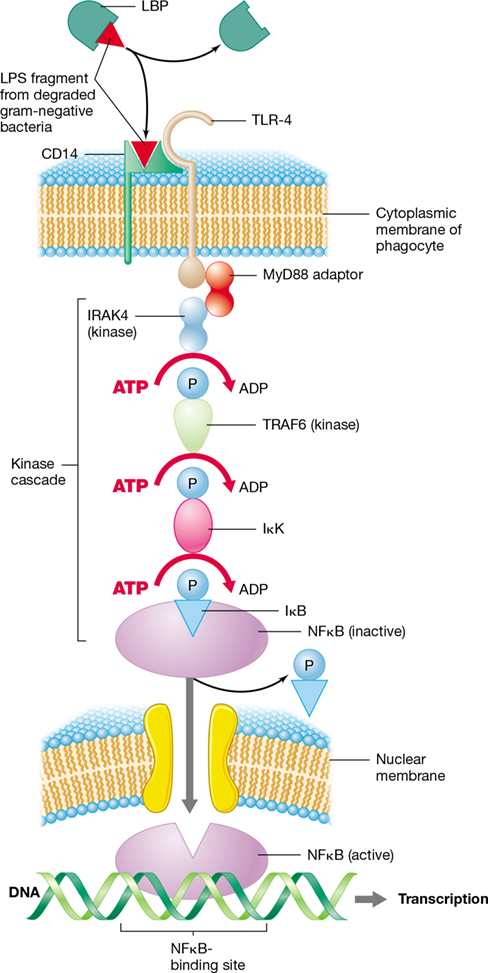
Figure 26.9 Signal transduction in innate immunity.

Signal transduction is the transmission of a molecular signal across a cytoplasmic membrane by way of chemical modifications—typically involving a series of proteins in a signaling cascade—that results in a response, such as differential gene expression. In innate immunity, signal transduction is initiated when LPS, a PAMP, is bound by LBP (lipopolysaccharide-binding protein), which then transfers LPS to CD14 on the surface of a phagocyte. The LPS–CD14 complex then binds to the transmembrane TLR-4 receptor. The binding of TLR-4 initiates a series of reactions involving adaptor proteins and kinases, resulting in activation of the transcription factor NFκ B. NFκ B then diffuses across the nuclear membrane, binds to DNA, and initiates transcription of genes encoding proteins essential for innate immunity.
TLR-4 is an integral protein having external, transmembrane, and cytoplasmic domains. The external domain of TLR-4 binds LPS complexed with a cell surface protein called CD14 (Figure 26.9). Binding of the CD14–LPS complex by the TLR-4 external domain causes a conformational change in TLR-4 that allows the cytoplasmic domain to interact with an adaptor protein called MyD88, which then binds a protein tyrosine kinase (PTK) called IRAK4. PTKs transfer energy-rich phosphates from ATP to target-protein tyrosines that are exposed when binding alters conformation. Binding of MyD88 by IRAK4 initiates a kinase cascade that triggers successive ATP-mediated phosphorylation of TRAF6, IκK (inhibitor of kappa kinase), and IκB (inhibitor of kappa B) proteins. Phosphorylation of IκB causes it to dissociate from, and thereby activate, NFκB. Activated NFκB can then diffuse across the nuclear membrane, bind to NFκB-binding motifs on DNA, and initiate transcription of downstream genes.
As Figure 26.9 shows, signal transduction pathways initiate activation of transcription through ligand–receptor binding on the surface of the phagocytic cell. The ligand–receptor interaction outside the cell induces the binding, recruitment, and concentration of the adaptor proteins and kinase enzymes inside the cell. A single kinase can phosphorylate many signal cascade proteins, thus amplifying the effect of a single ligand–receptor interaction. Signal transduction leading to activation of shared transcription factors and protein synthesis is also the mechanism by which lymphocytes are activated in adaptive immunity, as we discuss in Chapter 27.
Check Your Understanding
Identify a PAMP shared by a group of microorganisms. Then, identify the cell types that use PRRs to provide innate immunity to pathogens.
Outline the general features of a signal transduction pathway starting with binding of a PAMP by a membrane-associated PRR.
26.7 Phagocytosis and Phagocyte Inhibition
When phagocytes encounter infectious agents or their harmful products, the activation of signal transduction pathways (Figure 26.9) triggers genetic responses in the phagocyte that direct the containment and removal of the threat—phagocytosis (Figure 26.10). While these mechanisms effectively protect the body from most infectious exposures, they are not foolproof; many pathogens deploy effective defenses against phagocytes in an attempt to thwart the innate immune response.
Figure 26.10 Phagocytosis.

Time-lapse phase-contrast micrographs of the phagocytosis and digestion of a chain of Bacillus megaterium cells by a human phagocyte. The bacterial chain is about 20 μm long.
Phagocytosis and the Phagolysosome
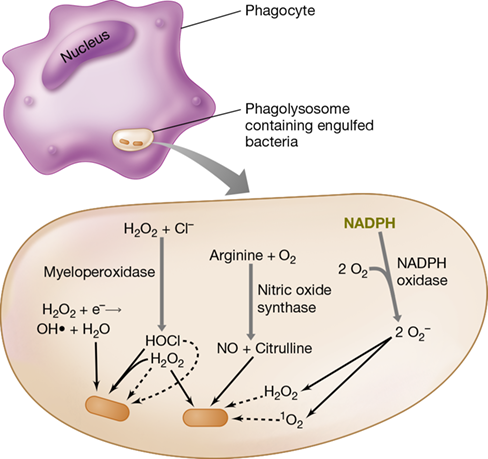
Most phagocytes contain multiple membrane-bound inclusions called lysosomes, cytoplasmic vesicles containing bactericidal substances, such as toxic oxygen compounds, lysozyme, proteases, phosphatases, nucleases, and lipases. Through the molecular mechanisms we have just discussed, phagocytes identify and engage pathogens on surfaces, such as blood vessel walls or fibrin clots, before initiating phagocytosis (Figure 26.10). Activation of the phagocyte through signal transduction causes the phagocyte membrane to envelop and engulf pathogens, eventually pinching off inwardly to form a phagosome, a vacuole containing the engulfed pathogen. The phagosome then moves into the cytoplasm and fuses with a lysosome to form a phagolysosome (Figure 26.11). The toxic chemicals and enzymes within the phagolysosome kill and digest the engulfed microbial cell.
Figure 26.11 Activity of phagocyte enzymes in generating toxic oxygen compounds.

These compounds include hydrogen peroxide (H2O2), the hydroxyl radical (OH⋅), hypochlorous acid (HOCl), the superoxide anion (O2−), singlet oxygen (1O2), and nitric oxide (NO). Formation of these toxic compounds requires a substantial increase in the uptake and utilization of molecular oxygen, O2. This increase in oxygen uptake and consumption by activated phagocytes is called the respiratory burst.
Genes that control the production of reactive oxygen compounds toxic to pathogens are highly transcribed in activated phagocytes. These reactive oxygen species include hydrogen peroxide (H2O2), superoxide anions (O2−), hydroxyl radicals (OH⋅), singlet oxygen (1O2), hypochlorous acid (HOCl), and nitric oxide (NO) (Figure 26.11; Section 4.16). Phagocytic cells use these toxic oxygen compounds to kill ingested bacterial cells by oxidizing key cellular constituents. The lethal oxidative reactions are contained within the phagolysosome, and this prevents damage to the phagocyte itself.
Inhibiting Phagocytes
Some pathogens have mechanisms for neutralizing toxic phagocyte products, for killing the phagocytes, or for avoiding phagocytosis. For example, several species of Mycobacterium produce pigmented compounds called carotenoids that neutralize singlet oxygen and prevent pathogen killing. In addition, Mycobacterium tuberculosis, the bacterium that causes tuberculosis, grows and persists within phagocytic cells (Section 31.4). Cells of M. tuberculosis use their cell wall glycolipids (Section 16.11) to absorb hydroxyl radicals and superoxide anions, the most lethal toxic oxygen species produced by phagocytes (Figure 26.11).
Some intracellular pathogens produce phagocyte-killing proteins called leukocidins. In such cases, the pathogen is ingested as usual, but the leukocidin kills the phagocyte, releasing the pathogen unharmed. Dead phagocytes make up much of the material of pus, and pyogenic (pus-forming) pathogens such as Streptococcus pyogenes (scarlet and rheumatic fevers) and Staphylococcus aureus (skin infections) are major leukocidin producers. Localized infections by pyogenic bacteria thus form boils or abscesses.
Another important pathogen defense against phagocytosis is the bacterial capsule (Section 2.6). Because the capsule prevents necessary molecular interactions between the surface of the phagocyte and that of the bacterial cell, encapsulated bacteria are often highly resistant to phagocytosis. For example, fewer than ten cells of an encapsulated strain of Streptococcus pneumoniae can kill a mouse within a few days (Section 25.3 and Figure 25.9), whereas strains lacking a capsule are harmless (Figure 1.38). Surface components other than capsules can also inhibit phagocytosis. For instance, pathogenic S. pyogenes produces M protein, a substance that alters the surface of the pathogen and inhibits phagocytosis.
Soluble PRRs and other host molecules such as antibodies (Chapter 27) can interact with capsules and other pathogen surface molecules, thereby reversing the protective effect of bacterial defense mechanisms and enhancing phagocytosis. As an example, the vaccine directed against Streptococcus pneumoniae, the agent of bacterial pneumonia, uses capsule polysaccharides to induce protective antibodies (Section 28.3). Thus, the battle between pathogen and the host innate immune system is a dynamic one, where both sides deploy multiple weapons in attempts to thwart the success of the other.
Check Your Understanding
Identify the mechanism used by phagocytes to induce pathogen killing.
Describe several reasons why phagocytes are not always effective at removing pathogens from the body.
IV Other Innate Host Defenses
In addition to phagocytes, weapons and defenses of the innate immune system include generalized responses, such as inflammation and fever, antibacterial proteins of the complement system, and antiviral natural killer cells and interferons.
In addition to the physical and chemical barriers to pathogen invasion and the direct destruction of pathogens by activated phagocytes, mammalian immune systems have other inborn mechanisms that help counter infection by pathogens. Although unpleasant for the host, inflammation and fever can be effective host defense strategies for controlling microbial growth in the body and eliminating pathogens. These mechanisms, along with a consideration of the complement system and the activities of cytotoxic natural killer cells, round out our discussion of the innate immune response.
26.8 Inflammation and Fever
Inflammation is a complex biological response to noxious stimuli, such as physical injury, toxins, and pathogens. Inflammation is characterized by redness (erythema), swelling (edema), pain, and heat, usually localized at the site of infection or injury (**Figure 26.12*a***). The mediators of inflammation are a group of cell activator and chemoattractant molecules, including cytokines and chemokines. Various cells, including those damaged by injury, produce these activators. The most important chemokines and cytokines are called proinflammatory because of their inflammation-inducing capacity, and they are produced in high concentrations by phagocytes and lymphocytes during pathogen challenge.
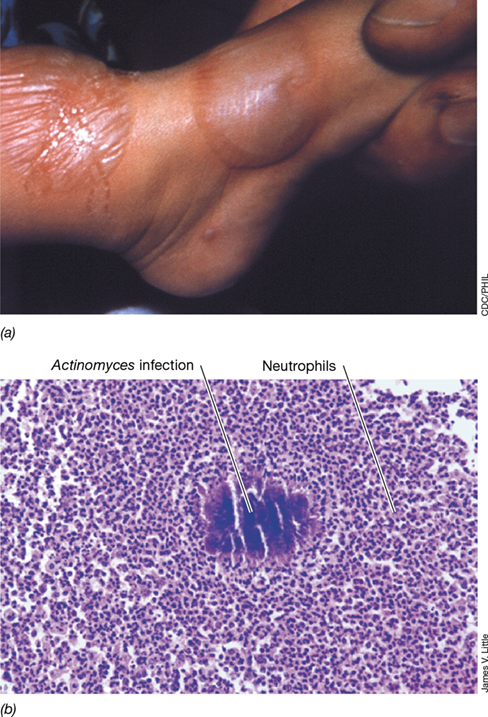
Figure 26.12 Inflammation.

(a) Photograph of a child’s foot showing swelling due to infection with vaccinia virus; fluid accumulation results from the inflammatory response. (b) Photomicrograph showing infection by Actinomyces, a filamentous bacterium. The stained cells surrounding the dark mass of bacteria in the center are neutrophils (Figure 26.5), indicating acute inflammation.
Both innate and adaptive immune responses to infection can cause inflammation, and both responses trigger the release of molecules that recruit and activate effector cells, such as neutrophils. Although it is a generalized immune response, inflammation plays a crucial role to isolate and limit tissue damage by initiating the destruction of pathogen invaders and removing damaged cells as tissues are repaired. As we will soon discuss, however, the inflammatory response may also inadvertently result in considerable damage to healthy host tissue.
Inflammatory Cells and Local Inflammation
Immune-mediated inflammation is an acute response that begins at the site of pathogen entry into the body. The PRRs on macrophages and other tissue cells at the site of infection engage the pathogen PAMPs (Figure 26.7). This activates local cells to produce and release mediators including cytokines and chemokines that interact with receptors on other cells, such as neutrophils (Figure 26.5). For example, local tissue macrophages that are activated by PAMP–PRR interaction secrete a chemokine called CXCL8. This molecule activates neutrophils to migrate along the chemokine gradient toward the source of the CXCL8, where they begin to ingest and kill the pathogen. The neutrophils, in turn, secrete even more CXCL8, attracting more neutrophils and amplifying the response, eventually destroying the pathogens (Figure 26.12b).
The chemokine and cytokine mediators released by injured cells and phagocytes contribute to inflammation. For example, macrophages and other cells at the site of infection produce proinflammatory cytokines including interleukin-1 (IL-1), IL-6, and tumor necrosis factor α(TNF-α). These cytokines increase vascular permeability, causing the edema, erythema, and local heating associated with inflammation (**Figure 26.13*a***). Although edema stimulates local neurons, causing pain, the pressure associated with swelling also serves to force fluids away from blood vessels and into the lymphatic system, simultaneously helping to strengthen the immune response and prevent the spread of pathogens to the bloodstream. If pathogens do enter the bloodstream, a condition called bacteremia, this could trigger the much more serious and potentially life-threatening septicemia (Section 25.2).
Figure 26.13 Local and systemic inflammation.

(a) Local infection, mediated by proinflammatory cytokines from local macrophages, results in inflammation that subsides as the infection is cleared. (b) Systemic infection causes systemic release of proinflammatory cytokines, resulting in widespread systemic inflammatory symptoms including severe edema, fever, and septic shock, even if the infection is controlled.
The usual outcome of the inflammatory response is a rapid localization and destruction of the pathogen by macrophages and recruited neutrophils. As the pathogens are destroyed, inflammatory cells are no longer stimulated, and as a result, their numbers at the infection site are diminished. As cytokine production decreases, the attraction of phagocytes to affected tissues lessens, and inflammation subsides.
Fever
Certain cytokines released during an inflammatory response induce fever, a condition of elevated body temperature. For example, the proinflammatory cytokines IL-1, IL-6, and TNF-α are endogenous pyrogens. These substances stimulate the hypothalamus, the temperature-controlling center of the brain, to produce prostaglandins, chemical signals that raise body temperature and cause fever. These same cytokines released in small amounts at local sites of infection induce localized heating, which increases blood flow and promotes healing. The release of endogenous pyrogens is a physiological response to the presence of exogenous pyrogens, components of pathogens that induce fever, such as the lipid A endotoxin of gram-negative bacteria LPS (Section 2.4).
Although uncomfortable for the host, fever is an important component of the innate immune response to some infections. The rise from normal human body temperature, about 37 °C (98.6°F), to fever temperature, usually 38–40 °C (100.4–104°F), is a beneficial response to infection because higher body temperatures inhibit growth of most pathogens. Slower growth limits multiplication of the invading pathogen, thereby minimizing tissue damage and easing the workload on immune cells, especially phagocytes. Elevated body temperatures also increase the production of transferrins, molecules that bind and sequester iron in blood and lymph, thus depriving pathogens of this important nutrient (Sections 3.1 and 25.4).
While low to moderate fever is an important component of the innate immune response, a continuous or uncontrolled rise in body temperature (>40 °C) is a rare but life-threatening condition that may accompany certain disease conditions and requires immediate medical intervention. This usually includes the administration of antipyretic (fever-reducing) medications, such as acetaminophen or ibuprofen, which counteract the effect of endogenous pyrogens on the hypothalamus.
Systemic Inflammation and Septic Shock
In some cases, the inflammatory response fails to localize the pathogens, and the reaction spreads throughout the body. Uncontrolled systemic inflammation can be more dangerous than the original infection, with inflammatory cells and mediators contributing to body-wide inflammation. An inflammatory response that spreads inflammatory cells and mediators through the entire circulatory and lymphatic systems can lead to septic shock, a life-threatening condition.
Although there are potentially many causes of pathogen-induced septic shock, one example is systemic infection by enteric bacteria, such as Salmonella species or Escherichia coli, which can be introduced into the peritoneal cavity or the bloodstream by a ruptured or leaking bowel. The primary infection is often cleared by the activity of phagocytes and antibiotic treatment. However, the endotoxic outer membrane LPS from these gram-negative bacteria interacts with a PRR on phagocytes, stimulating production of proinflammatory cytokines that are released into the circulation. These cytokines induce systemic responses that parallel the localized inflammatory response but on a much larger scale that affects many organ systems, ultimately leading to a body-wide inflammatory event (Figure 26.13b). The result is a massive efflux of fluids from the central vascular tissue, causing a loss of systemic blood pressure and the influx of fluids from vascular tissues into extravascular spaces. Septic shock causes death in up to 30% of affected individuals.
Check Your Understanding
Identify the molecular mediators of inflammation and fever and define their individual roles.
Identify the major symptoms of localized inflammation and of septic shock.
26.9 The Complement System
The complement system, or simply complement, is a group of sequentially interacting plasma proteins, many with enzymatic activity, that functions to boost the efficiency of both innate and adaptive immune responses for the destruction of pathogens. Found throughout the body, complement proteins are produced in the liver and can be activated by the presence of a pathogen. Upon pathogen recognition, complement proteins interact with one another to trigger a cascade of events in a process called complement activation, the major outcomes of which are enhanced phagocytosis, inflammation, and lysis of invading cells.
The individual proteins of complement are designated as C (for complement) followed by a number or letter for the subunits, as C1, C2, C3, and so on. Complement proteins exist in an inactive conformation until they are enzymatically split to assume their active forms. The cleaving of C3 to its products, C3a and C3b, is the key event in complement activation. Three different mechanisms (pathways) are known that lead to complement activation, and we consider each of these now.
Classical Complement Activation
The classical pathway of complement activation is initiated when complement proteins interact with antibodies bound to pathogen surfaces. The antibodies are said to fix (bind) the ever-present complement proteins, and thus, classical complement activation depends upon the adaptive immune response. The complement proteins react in a defined sequence, or cascade, with activation of one complement protein leading to activation of the next, and so on.
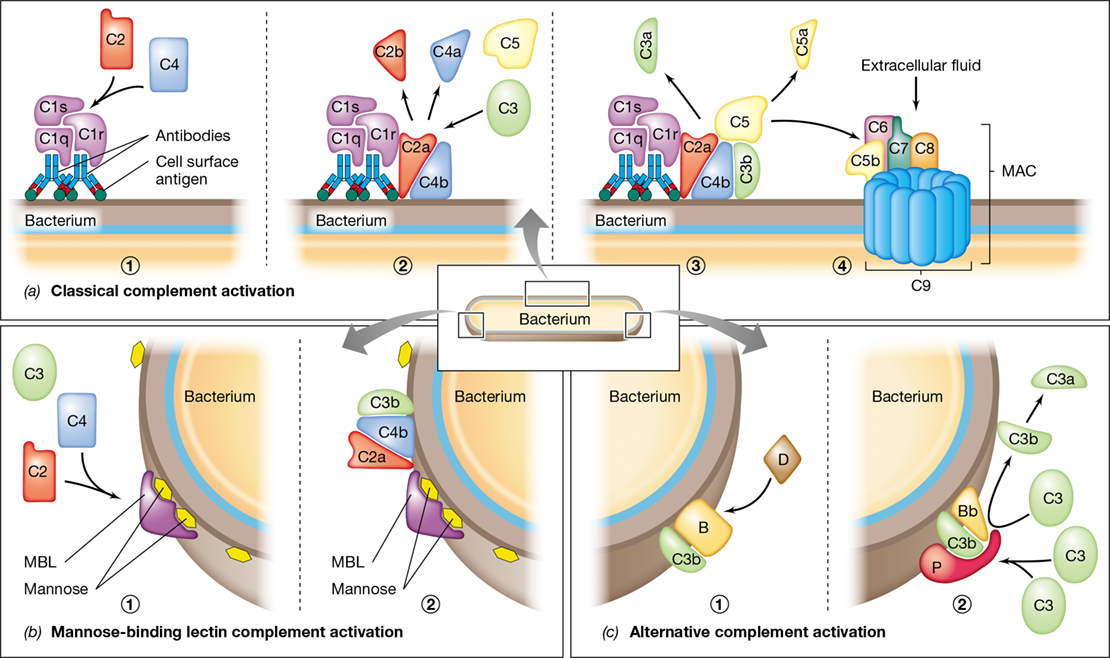
The key steps of classical activation of complement, shown in **Figure 26.14*a***, start with binding of antibody to antigen (initiation), followed by binding of the C1 complex (C1q, C1r, and C1s) to the antibody–antigen complex. This complex recruits and cleaves C2 into its fragments, C2a and C2b, and C4 into its fragments, C4a and C4b. C2a and C4b interact and are deposited on the pathogen surface. The resulting C2a-C4b complex functions as a C3 convertase, an enzyme that cleaves C3 to yield C3a and C3b. C3b then binds to the convertase, forming a C5 convertase that cleaves C5 into C5a and C5b. The liberated C5b then binds C6 and C7, forming a complex that inserts into the membrane of the pathogen. The C5b–7 complex recruits C8 and several copies of C9, forming a large C5b–9 unit called the membrane attack complex (MAC). The MAC forms a pore through the cytoplasmic membrane of the pathogenic cell, allowing extracellular fluids to rush in and lyse the cell (Figure 26.14a; see also Figure 26.15b). Dozens to hundreds of MACs may perforate a single bacterial cell at the point of lysis.
Figure 26.14 Complement proteins and complement activation in the immune response.

(a) The sequence, orientation, and activity of the components of the classical complement pathway as they interact to lyse a cell. 1 Binding of the antibody and the C1 protein complex (C1q, C1r, and C1s). 2 The C2a-C4b complex is a C3 convertase that splits C3. 3 The C2a-C4b-C3b complex cleaves C5, and C5b then binds an adjacent membrane site. 4 Sequential binding of C6, C7, C8, and C9 to C5b produces a pore, the membrane attack complex (MAC), in the membrane. (b) The mannose-binding lectin (MBL) pathway. 1 MBL binds to mannose on the bacterial surface and recruits C2 and C4. 2 MBL anchors formation of C2a-C4b-C3b. This complex activates C5, as in step 3 of part a, and initiates formation of the MAC (step 4 in a). (c) The alternative pathway. 1 C3b bound to the cell binds protein B, which is cleaved by protein D. 2 The resulting C3b-Bb complex is stabilized on the membrane by factor P (properdin) and then acts on C3 in the blood, causing more C3b to bind to the membrane. Bound C3b-Bb-P then activates C5, as in step 3 of the classical activation pathway above, and initiates formation of the MAC (step 4 in a).
When activated by specific antibody, MAC formation lyses many gram-negative bacteria. However, gram-positive bacteria, such as Streptococcus species, are not usually lysed by complement because their thick cell walls make the cytoplasmic membrane less accessible to MAC proteins. Gram-positive bacteria can, however, be destroyed through opsonization.
Opsonization is the coating of pathogens with host immune proteins, such as antibodies or C3b, resulting in enhanced phagocytosis of target cells (**Figure 26.15*a***). Opsonization neutralizes pathogens and makes them much more likely to be identified, engulfed, and destroyed by phagocytes. This is because most phagocytes, including neutrophils and macrophages, have antibody receptors (FcR) and C3b receptors (C3R) on their surfaces, which bind antibody and C3b complement protein, respectively. Normal phagocytic processes are enhanced about 10-fold by antibody–FcR interactions and amplified another 10-fold by C3b–C3R interactions.
Figure 26.15 Complement proteins and the outcomes of complement activation.

Each complement activation pathway leads to the production of a C3 convertase that cleaves C3 to its active products, C3a and C3b. These initiate three possible outcomes: opsonization, lysis by membrane attack complex (MAC) formation, and inflammation. (a) Pathogens coated with C3b (green) or with specific antibodies become opsonized, allowing for enhanced recognition by phagocytes through C3R and FcR proteins. (b) Transmembrane pores (white arrows) formed by MAC components C5b through C9 cause lysis of target cells. The transmission electron micrograph is a negative stain image showing human MACs attacking a foreign (rabbit) erythrocyte. (c) During complement-activated inflammation, liberated C3a and C5a bind receptor proteins on mast cells, causing the mast cells to degranulate and release proinflammatory histamine.
By-products of complement activation include chemoattractants called anaphylatoxins; these molecules cause inflammatory reactions at the site of complement deposition. For example, when C3 is cleaved to C3a and C3b, C3b opsonizes the target cell as described above; meanwhile, release of soluble C3a attracts and activates phagocytes, increasing phagocytosis. In addition, both C3a and the C5a cleavage product are able to bind receptors on the surface of mast cells, causing them to degranulate and release large amounts of proinflammatory histamine (Figure 26.15c).
Mannose-Binding Lectin and Alternative Pathways
The mannose-binding lectin pathway and alternative pathway of complement activation (Figure 26.14b and c) depend on recognition of shared pathogen components by complement proteins and are an important part of the innate immune response. The mannose-binding lectin (MBL) pathway depends on the activity of a serum MBL protein. MBL is a soluble PRR (Section 26.6) that binds to mannose-containing polysaccharides found only on bacterial cell surfaces (Figure 26.14b). The MBL–polysaccharide complex is similar to the C1 complex of the classical pathway (Figure 26.14a) in that it fixes C2a and C4b, again producing C3 convertase and binding C3b to C2a-C4b. As before, this complex catalyzes formation of the C5–9 MAC and leads to lysis or opsonization of the bacterial cell.
The alternative pathway is a nonspecific complement activation mechanism that uses many of the classical complement pathway components, as well as several unique serum proteins. Together they induce pathogen opsonization and phagocytosis and activate the C5–9 MAC. C3b is produced by spontaneous hydrolysis of C3 into C3a and C3b, which occurs at low levels in the blood and tissues at all times. If pathogens such as bacteria are present, C3b binds to the bacterial cell surface (opsonization). C3b can then bind the alternative pathway serum protein factor B, which is cleaved by factor D to yield soluble Ba and Bb (Figure 26.14c). The C3b-Bb complex may then be bound by factor P (properdin) to form C3b-Bb-P, another C3 convertase. C3b-Bb-P then attracts and cleaves additional C3, depositing more C3b on the membrane and amplifying the remaining steps of the complement cascade (Figure 26.15).
Both the alternative pathway and the MBL pathway nonspecifically target bacterial invaders and lead to activation of opsonization (Figure 26.15a) and MACs (Figure 26.15b) via formation of stable C3 convertases. MBL, factors B, D, and P, and classical complement proteins are part of the innate immune response, but unlike the classical pathway, neither the alternative pathway nor the MBL pathway requires prior antigen exposure or the presence of antibodies for activation.
We now move on to consider the immune system’s response to viruses rather than bacterial pathogens. Many pathogens are viruses, and because of their small size and unique properties, special immune defense mechanisms are required to deal with them.
Check Your Understanding
In what ways does the classical pathway of complement activation differ from the mannose-binding lectin and alternative pathways?
What is opsonization, and how does opsonization help fight bacterial infection?
Why are the mannose-binding lectin and alternative pathways considered part of the innate immune system?
26.10 Innate Defenses Against Viruses
In addition to opsonization and the activity of phagocytes, the immune system has other innate defenses that are especially important for controlling and eliminating viral infections. These include natural killer cells (Figure 26.5) and interferons. Natural killer cells are lymphocyte-like cells that recognize and kill compromised (unhealthy) cells, such as those infected with intracellular pathogens, in particular viruses. Whereas natural killer cells help rid the body of already infected cells, interferons, small proteins of the cytokine family, help healthy cells ward off viral infection. We consider both of these innate defenses here.
Natural Killer Cells
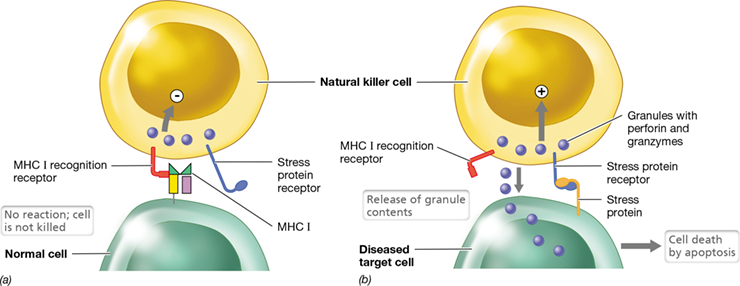
Natural killer cells (NK cells) are cytotoxic innate-immunity lymphoid cells that lack BCRs and TCRs and, thus, are distinct from T cells and B cells (Figure 26.5). The role of NK cells is to seek out and destroy compromised cells, such as cells infected with intracellular pathogens (such as viruses) or cancer cells (Figure 26.16). When an NK cell engages an infected or otherwise diseased cell, granules in the cytoplasm of the NK cell migrate to the contact site and release their contents against the target cell. These granules contain perforin and proteases called granzymes. Perforin binds the membrane of the target cell and forms a pore through which granzymes enter the target cell (Figure 26.16b). Granzymes are cytotoxins that cause apoptosis, or programmed cell death, characterized by non-inflammatory DNA degradation and membrane blebbing. During the killing process, NK cells remain unaffected and their membranes are not damaged by perforin. Also during this process, NK cell numbers do not increase nor do NK cells exhibit immune memory after interaction with target cells. With this in mind, how do NK cells recognize compromised host cells that should be destroyed? The key to this mechanism is the heterodimeric membrane protein found on the surface of most cells: the major histocompatibility complex (MHC) protein.
Figure 26.16 Natural killer cells.

Natural killer (NK) cells have two receptors: One interacts with MHC I on healthy cells; the second receptor interacts with cell stress proteins, found only on tumor cells or pathogen-infected cells. (a) Recognition of MHC I on healthy cells prevents the NK cell from releasing its toxic contents. (b) Pathogen-infected cells or tumor cells express stress proteins and often reduce MHC I expression. Especially in the absence of MHC I recognition, the NK cell interacts with the stress protein and releases perforin and granzymes, inducing apoptosis and killing the diseased cell.
Mastering Microbiology
There are two classes of MHC proteins—class I and class II—and their primary function is to present antigen to various immune cells to trigger an immune response (Chapter 27). MHC II proteins are expressed on antigen-presenting cells (APCs) only, which include B cells, macrophages, and dendritic cells (Figure 26.5). By contrast, MHC I proteins are found on the surfaces of all nucleated cells. In uninfected cells, which contain no pathogens or foreign antigens, MHC I proteins bind and present self peptides, protein fragments derived from the normal degradation of self proteins, to the external environment (the term “self” here means “native to the cell”). In cells that have been infected by viruses or other intracellular pathogens, MHC I proteins display peptides derived from the infectious agent. This serves as a signal to a special type of T lymphocyte called a T-cytotoxic cell to destroy the infected cell.
Because MHC I proteins facilitate the destruction of cells infected with an intracellular pathogen, such as a virus, many viral genomes encode proteins that suppress expression of MHC I in host cells. Without MHC I proteins on their surfaces—a condition that is also common in cancer cells—T-cytotoxic cells cannot identify and eliminate diseased host cells. This, then, is where NK cells play a major role; NK cells recognize and destroy compromised host cells that have reduced expression of MHC I on their surfaces.
NK cells recognize and destroy pathogen-infected or tumor cells by using a two-receptor system. The molecular targets of NK cells are MHC I proteins on the surface of other cells (Figure 26.16a). As NK cells circulate and interact with other cells in the body, they use special receptors on their surfaces to recognize MHC I proteins on normal, healthy cells. Binding of the MHC I recognition receptors on NK cells to MHC I on other cells deactivates the NK cell, turning off the perforin and granzyme killing mechanisms. In addition to a deficiency of MHC I proteins, pathogen-infected or tumor cells frequently express stress proteins on their surfaces; NK cells have complementary receptors for many of these stress proteins. Especially in the absence of the MHC I interactions, the stress receptors on NK cells engage stress proteins on target cells. This interaction triggers the release of perforin and granzymes from the NK cell (Figure 26.16b). In this way, pathogen-infected or tumor cells that exhibit stress proteins and no longer express the MHC I proteins of healthy cells are removed from the body.
Mastering Microbiology
Art Activity: Figure 26.16 Antiviral activity of interferons
In addition to these functions, NK cells are the primary effector cells of antibody-dependent cell-mediated cytotoxicity (ADCC), a process by which cells coated (opsonized) with a class of antibody called immunoglobulin G (IgG; Section 27.3) are lysed by cytotoxic immune cells. Using special receptors on their surfaces, NK cells engage IgG antibodies that have previously bound a target cell. This interaction activates the NK cell, causing it to release its cytotoxic contents and kill the opsonized target cell.
Interferons
Interferons, in particular IFN-α and IFN-β, are small proteins in the cytokine family that prevent viral replication by stimulating the production of antiviral proteins in uninfected cells (Figure 26.17). Host cells produce and secrete interferons in response to certain viral infections or exposure to inactivated viruses or viral nucleic acids. Interferons are produced in large amounts by cells infected with viruses of low virulence, but highly virulent viruses inhibit host protein synthesis before interferon can be produced, significantly reducing interferon production. The presence of double-stranded RNA (dsRNA) also induces interferon synthesis. In nature, dsRNA exists only in certain RNA viruses, such as rhinoviruses (one of many common cold viruses, Section 31.7); the viral dsRNA stimulates the animal cell to synthesize and release interferon.
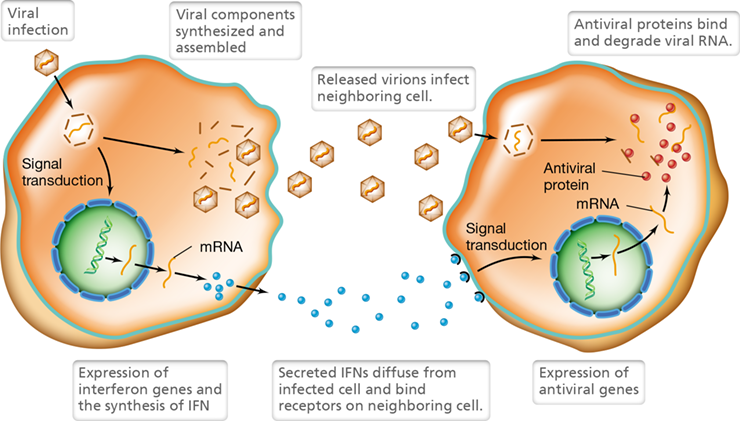
Figure 26.17 Antiviral activity of interferons.

Host cells secrete interferons, a type of cytokine, in response to viral infection. The interferons bind to uninfected cells, triggering a signal transduction pathway that leads to the synthesis of proteins that bind viral nucleic acids and interfere with viral replication.
Antiviral interferon activities are host-specific rather than virus-specific. That is, interferon produced by a species activates receptors only on cells from the same species. As a result, interferon produced by cells of an animal in response to, for example, a rhinovirus, could also inhibit multiplication of, for example, influenza viruses in cells of the same species. However, the interferon would have no effect on the multiplication of viruses, including the original rhinovirus, in other species. We examine the potential use of antiviral interferons as chemotherapeutic or prophylactic treatments in Chapter 28.
A third type of interferon, IFN-γ, stimulates phagocytes and is produced by NK cells in response to cytokine signaling. For example, macrophages that have ingested microorganisms produce the cytokine interleukin-12 (IL-12). NK cells receive the IL-12 signal and become activated, causing them to secrete IFN-γ. The resulting exposure to IFN-γ activates the macrophages and causes them to more efficiently kill ingested microbes. As we will see in the next chapter, similar cooperation occurs between antigen-presenting phagocytes and a subset of T lymphocyte called a T-helper cell (Section 27.8), and this interaction serves as a key link between innate and adaptive immunity.
Check Your Understanding
Identify and compare the targets and the recognition mechanisms used by T-cytotoxic cells and NK cells.
Under what conditions are interferons produced, and how do they limit the transmission of viruses from one host cell to another?
Explore the Microbial World Pattern Recognition Receptors of Hydrothermal Vent Tube Worms Facilitate Endosymbiosis
Explore the Microbial World Pattern Recognition Receptors of Hydrothermal Vent Tube Worms Facilitate Endosymbiosis
Pattern Recognition Receptors of Hydrothermal Vent Tube Worms Facilitate Endosymbiosis
Invertebrates and plants lack adaptive immunity but have a well-developed innate immune response to a wide variety of pathogens. As discussed in Section 26.6, virtually all multicellular organisms respond to pathogen invasion by recognizing signature molecules found on pathogen surfaces. These molecules contain conserved, repetitive structures called pathogen-associated molecular patterns (PAMPs) that include molecules such as the lipopolysaccharide (LPS) and flagellin of gram-negative bacteria, the peptidoglycan of gram-positive bacteria, and the double-stranded RNA of certain viruses. The term microbe-associated molecular pattern, or MAMP, is also commonly used to describe signature molecules found on microorganisms. However, “MAMP” is a broader term than “PAMP” because it includes components found on microorganisms that are not pathogenic.
Unlike PAMPs, which are exploited specifically for innate defense against pathogens, MAMPs found on nonpathogenic bacteria can serve an entirely different purpose—that of facilitating, through host pattern recognition receptors (PRRs), a mutualistic partnership rather than a confrontation between a host and the bacteria that colonize it. As we learned in Chapter 23, a wide variety of plants and animals maintain symbiotic relationships with microorganisms. There we discussed the association of tube worms that develop near hydrothermal vents in the deep sea with autotrophic, sulfur-oxidizing bacteria (SOB) that inhabit their trophosome, a spongy internal organ that comprises most of the volume of the 1- to 2-m-long worms (Figure 1). These SOB form an endosymbiosis with the worms in which the bacteria provide all organic carbon requirements for their animal host in exchange for a steady supply of essential metabolites, in particular H2S, O2, and CO2. H2S is the energy source for the SOB; they oxidize it to S0 and then SO4 2− and respire the electrons to generate a proton motive force that drives ATP synthesis. Oxygen (O2) is required as a terminal acceptor of electrons that have traversed the electron transport chain. CO2 is the carbon source and is incorporated into bacterial cell material by way of the Calvin cycle, the major means of autotrophy in chemolithotrophic bacteria.
Figure 1 Hydrothermal vent tube worms harboring endosymbiotic sulfur-oxidizing bacteria.
Top: A “black smoker” hydrothermal vent community containing several tube worms that obtain organic carbon from sulfur-oxidizing chemolithotrophic bacteria (SOB) living within them. Bottom: A close-up view of tube worms; each worm is 1–2 m long. The red area on the top of each worm, called the plume, is where O2 and H2S are taken in to be fed to the worm’s SOB endosymbionts residing in the trophosome.
U.S. National Oceanic and Atmospheric Administration
This fascinating association raises the question of how it is established. Specifically, how does the tube worm populate its trophosome with SOB to the exclusion of other, potentially pathogenic, bacteria? The answer appears to be closely linked to MAMPs associated with the endosymbiotic SOB. Although host PRRs are typically used to recognize and eliminate pathogens, the study of tube worms and other animals that harbor endosymbiotic microbes shows a broader functionality for PRRs in that they can also interact beneficially with MAMPs to selectively populate a host with nonpathogenic symbionts.
The tube worm trophosome contains a large number of specialized host immune cells called bacteriocytes, and it is within these cells that the bacterial symbionts take up residence. The tube-worm bacteriocytes express high levels of PRRs that recognize MAMPs, such as specific cell surface lipoproteins associated with SOB. This positive interaction locates the bacteria to the trophosome and, with a steady supply of simple nutrients from the hydrothermal vent system delivered by blood circulating in the worm, stimulates colonization and growth of the symbionts in their animal host.
As this example illustrates, in addition to providing a rapid response to pathogen challenge, innate immune mechanisms—specifically, the interaction of PRRs with MAMPs—may also serve the primary role in governing host–symbiont interactions and the establishment of endosymbiotic relationships. Given the critical role innate immune mechanisms play in maintaining the animal–bacterial symbiosis within the tube-worm trophosome, it is likely that similar tightly choreographed molecular mechanisms constitute a host–microbe “dialogue” that helps to maintain balanced communities of beneficial microbiota in virtually all animals, including humans.
Chapter Review
Go to Mastering Microbiology for videos, animations, practice tests, and more.
I Fundamentals of Host Defense
26.1 Innate immunity is an inborn protective response to infection characterized in part by recognition and elimination of common pathogens, primarily through the activity of phagocytes. Adaptive immunity is the acquired ability of the immune system to eliminate specific pathogens from the body via lymphocyte-mediated responses, including the production of antibodies that bind foreign antigens on pathogens or their products.
Q Compare and contrast the major features of innate immunity with those of adaptive immunity.
26.2 The human body possesses numerous protective defenses against infectious agents. Natural host resistance to infection includes physical barriers posed by the skin and mucosa, as well as chemical barriers including acidic secretions, defensins, and lysozyme. The specificity of pathogens for particular tissues limits which hosts and tissues might be susceptible to infection.
Q Identify at least four mechanisms by which a healthy host resists infection.
II Cells and Organs of the Immune System
26.3 Cells involved in innate and adaptive immunity originate from hematopoietic stem cells in bone marrow. The blood and lymph systems circulate cells and proteins that are important components of the immune response. Diverse leukocytes participate in immune responses in all parts of the body.
Q Describe the significance of bone marrow, blood, and lymph to cells and proteins associated with the immune system.
26.4 Leukocytes are differentiated white blood cells derived from either myeloid or lymphoid precursor cells. Cells of myeloid lineage include monocytes and granulocytes. Monocytes include macrophages and dendritic cells, specialized phagocytes that function as antigen-presenting cells (APCs) to initiate an adaptive immune response. Granulocytes include neutrophils and eosinophils, which are phagocytes but not APCs, and basophils and mast cells, which are important inducers of the inflammatory response but may also cause allergic reactions. Lymphocytes include B and T cells, which facilitate adaptive immunity. Natural killer (NK) cells, which play a key role in destroying virus-infected and cancerous host cells, are also derived from a lymphoid precursor.
Q What is the origin of the phagocytes and lymphocytes active in the immune response?
III Phagocyte Response Mechanisms
26.5 Pathogens may colonize host tissues when appropriate nutrients and growth conditions are present, such as on mucosal surfaces, especially where the composition of the normal microbiota has been altered. Innate responses to microbial invasion and tissue damage are initiated by the release of chemokines, which recruit phagocytes and other immune cells to sites of infection.
Q Describe a scenario in which microorganisms invade body tissues. What factors allow for the migration of phagocytes to sites of infection?
26.6 Common pathogens carry pathogen-associated molecular patterns (PAMPs) on their surfaces and intracellularly. Phagocytes recognize PAMPs through preformed (innate) pattern recognition receptors (PRRs). The recognition and interaction process stimulates phagocytes to destroy the pathogens through a signal transduction mechanism that induces phagocytosis of the infectious agent.
Q Identify some PAMPs that are recognized by PRRs. Which cells express PRRs? How do PRRs associate with PAMPs to promote innate immunity?
26.7 Phagocytosis is the engulfing of infectious particles by phagocytes. Engulfed pathogens are bathed in toxic oxygen compounds and enzymes inside the phagolysosome, killing and degrading them. However, some pathogens have developed various defense mechanisms to avoid or inhibit phagocytes, including secretion of leukocidins, the presence of a capsule, and biosynthesis of carotenoid pigments, which combat oxidative stress.
Q Explain how phagocytes kill microorganisms, with particular attention to oxygen-dependent mechanisms. Then identify at least three properties of pathogens that inhibit the effectiveness of phagocytes.
IV Other Innate Host Defenses
26.8 Fever and inflammation, characterized by pain, swelling (edema), redness (erythema), and heat, are normal and generally beneficial outcomes that result from activation of nonspecific immune response effectors. However, uncontrolled systemic inflammation—called septic shock—or excessively high fever can lead to serious illness or death.
Q Identify the cells that initiate inflammation and the cells that are activated by inflammatory signals.
26.9 The complement system is composed of soluble proteins that catalyze bacterial opsonization and cell lysis. Complement is triggered by antibody interactions or by interactions with nonspecific activators, such as mannose-binding lectin. Complement activation may occur through either innate or antibody-dependent mechanisms. Complement may enhance phagocytosis, cause target cell lysis, or induce an inflammatory response.
Q Describe the complement system. Is the order of protein interactions important? Why or why not? Identify the components of the mannose-binding lectin pathway for complement activation. Identify the components of the alternative pathway for complement activation. How do these complement activation pathways differ from the classical pathway?
26.10 Through an antigen-independent mechanism, natural killer cells respond to both the presence of stress proteins and the absence of major histocompatibility complex (MHC) I on the surface of virus-infected cells or tumor cells, using perforin and granzymes to kill the compromised target cells. Interferons are cytokines produced by virus-infected cells that limit the spread of infection by stimulating the expression of antiviral proteins in uninfected cells. Other interferons activate phagocytes to enhance phagocytosis.
Q What is the activation signal for NK cells? How does this differ from the activity of T-cytotoxic lymphocytes?
Application Questions
Describe the relative importance of innate immunity compared to adaptive immunity. Is one more important than the other? Could humans survive in a normal environment without an immune system?
Describe the potential problems that would arise if a person had an acquired inability to phagocytose pathogens. Could the person survive in a normal environment such as a college campus? What defects in the phagocyte might cause lack of phagocytosis? Explain.
Inflammation is the hallmark of an active immune response. Explain how inflammation is triggered by innate immune mechanisms. Why does inflammation subside as an infection is controlled?
Do you agree with the following statement? Complement is a critical component of antibody-mediated defense. Explain your answer. What might happen to persons who lack complement component C3? C5? Factor B (alternative pathway)? Mannose-binding lectin (MBL)?
Chapter Glossary
the acquired ability to recognize and destroy a particular pathogen or its products, dependent on previous exposure to the pathogen or its products Antibody
a soluble protein produced by B cells and plasma cells that interacts with antigen; also called immunoglobulin Antibody-dependent cell-mediated cytotoxicity (ADCC)
the process by which NK cells bind IgG-coated target cells and kill them by secreting cytolytic toxins Antigen
a molecule capable of interacting with specific components of the immune system and that often functions as an immunogen to elicit an adaptive immune response Antigen-presenting cell (APC)
a macrophage, dendritic cell, or B cell that takes up and processes antigen and presents it to T-helper cells Basophil
a circulating granulocyte that contains granules of inflammatory mediators that, in some cases, contribute to allergic reactions B lymphocyte (B cell)
a lymphocyte that has immunoglobulin surface receptors, produces immunoglobulin, and may present antigens to T cells B cell receptor (BCR)
a cell surface immunoglobulin that acts as an antigen receptor on a B cell Chemokine
a soluble protein that recruits immune cells to an injury site; a type of cytokine Complement system
a series of proteins that react sequentially with antibody–antigen complexes, mannose-binding lectin, or alternative activation pathway proteins to amplify or potentiate target cell destruction Cytokine
a soluble protein produced by a leukocyte or damaged host cell; modulates an immune response Dendritic cell
a phagocytic antigen-presenting cell found in various body tissues; transports antigen to secondary lymphoid organs Epitope
the portion of an antigen that reacts with a specific antibody or T cell receptor Eosinophil
a phagocytic granulocyte especially active against extracellular parasites, such as helminths Fever
an increase in body temperature resulting from infection or the presence of toxins in the body Granulocyte
a leukocyte derived from a myeloid precursor that contains cytoplasmic granules consisting of toxins or enzymes that are released to destroy target cells Hematopoiesis
the process by which precursor stem cells in the bone marrow or gut differentiate into any type of blood cell Hematopoietic stem cell
a progenitor cell found primarily in the bone marrow but also in the gut that can differentiate into any of a variety of blood cells Immune memory (memory)
the ability to rapidly produce large quantities of specific immune cells or antibodies after subsequent exposure to a previously encountered antigen Immunity
the ability of an organism to resist infection Immunoglobulin
a soluble protein produced by B cells and plasma cells that interacts with antigen; also called antibody Inflammasome
a cytosolic signaling complex that contains NOD-like receptor pyrin 3 (NLRP3) and mediates the activation of potent proinflammatory cytokines Inflammation
a nonspecific reaction to noxious stimuli such as toxins and pathogens, characterized by redness (erythema), swelling (edema), pain, and heat, usually localized at the site of infection Innate immunity
the inherent ability to recognize and destroy an individual pathogen or its products that does not rely on previous exposure to a pathogen or its products Interferons
cytokine proteins produced by virus-infected cells that induce signal transduction in nearby cells, resulting in transcription of antiviral genes and expression of antiviral proteins Invasion
the ability of a pathogen to enter into host cells or tissues, spread, and cause disease Leukocyte
a nucleated cell in blood; also called a white blood cell Lymph nodes
organs that contain lymphocytes and phagocytes arranged to encounter microorganisms and antigens as they travel through the lymphatic circulation Lymphocytes
a subset of nucleated cells in blood involved in the adaptive immune response; B cells and T cells Macrophage
a large leukocyte found in tissues that has phagocytic and antigen-presenting capabilities Major histocompatibility complex (MHC)
a genetic region that encodes several proteins important for antigen processing and presentation; MHC I proteins are expressed on all nucleated cells, whereas MHC II proteins are expressed only on antigen-presenting cells Mast cell
tissue granulocyte similar in function to basophils that contains intracytoplasmic granules with inflammatory mediators Monocyte
Circulating phagocyte that contains many lysosomes and can differentiate into a macrophage or dendritic cell Mucosa-associated lymphoid tissue (MALT)
a part of the lymphatic system that interacts with antigens and microorganisms that enter the body through mucous membranes, including those of the gut, the genitourinary tract, and the respiratory tract Natural killer (NK) cell
a specialized lymphocyte that recognizes and destroys infected host cells or cancer cells in a nonspecific manner Neutrophil
a leukocyte exhibiting phagocytic properties, a granular cytoplasm (granulocyte), and a multilobed nucleus; also called polymorphonuclear leukocyte or PMN Opsonization
the deposition of antibody or complement protein on the surface of a pathogen or other antigen that results in enhanced phagocytosis Pathogen-associated molecular pattern (PAMP)
a repeating structural component of a microorganism or virus recognized by a pattern recognition receptor (PRR) Pattern recognition receptor (PRR)
a protein that is soluble or bound to a phagocyte membrane and that recognizes a pathogen-associated molecular pattern (PAMP) Phagocyte
a cell that engulfs foreign particles, and can ingest, kill, and digest most pathogens Phagocytosis
a mechanism for ingesting particulate material in which a portion of the cytoplasmic membrane surrounds the particle and brings it into the cell; for phagocytes of the immune system, the process of engulfing and killing foreign particles and cells Phagosome
an intracytoplasmic vacuole containing engulfed materials, especially pathogens or foreign particles Plasma
the liquid portion of the blood containing proteins and other solutes Plasma cell
a differentiated B cell that produces soluble antibodies Primary lymphoid organ
an organ in which antigen-reactive lymphocytes develop and become functional; the bone marrow is the primary lymphoid organ for B cells; the thymus is the primary lymphoid organ for T cells Secondary lymphoid organ
an organ at which antigens interact with antigen-presenting phagocytes and lymphocytes to generate an adaptive immune response; these include lymph nodes, spleen, and mucosa-associated lymphoid tissue (MALT) Serum
the liquid (noncellular) portion of the blood with clotting proteins removed Specificity
the ability of cells of the adaptive immune response to interact with particular antigens T lymphocytes (T cells)
a lymphocyte that interacts with antigens through a T cell receptor for antigen; T cells are divided into functional subsets including Tc (T-cytotoxic) cells and Th (T-helper) cells. Th cells are further subdivided into Th1 (inflammatory) cells and Th2 cells, which aid B cells in antibody formation T cell receptor (TCR)
an antigen-specific receptor protein on the surface of T cells Toll-like receptor (TLR)
one of a family of pattern recognition receptors (PRRs) found on phagocytes, structurally and functionally related to Toll receptors in Drosophila, that recognize a pathogen-associated molecular pattern (PAMP)